

Protracted yet Coordinated Differentiation of Long-Lived SARS-CoV-2-Specific CD8+ T Cells during Convalescence
- PMID: 34389625
- PMCID: PMC8763019
- DOI: 10.4049/jimmunol.2100465
Protracted yet Coordinated Differentiation of Long-Lived SARS-CoV-2-Specific CD8+ T Cells during Convalescence
Abstract
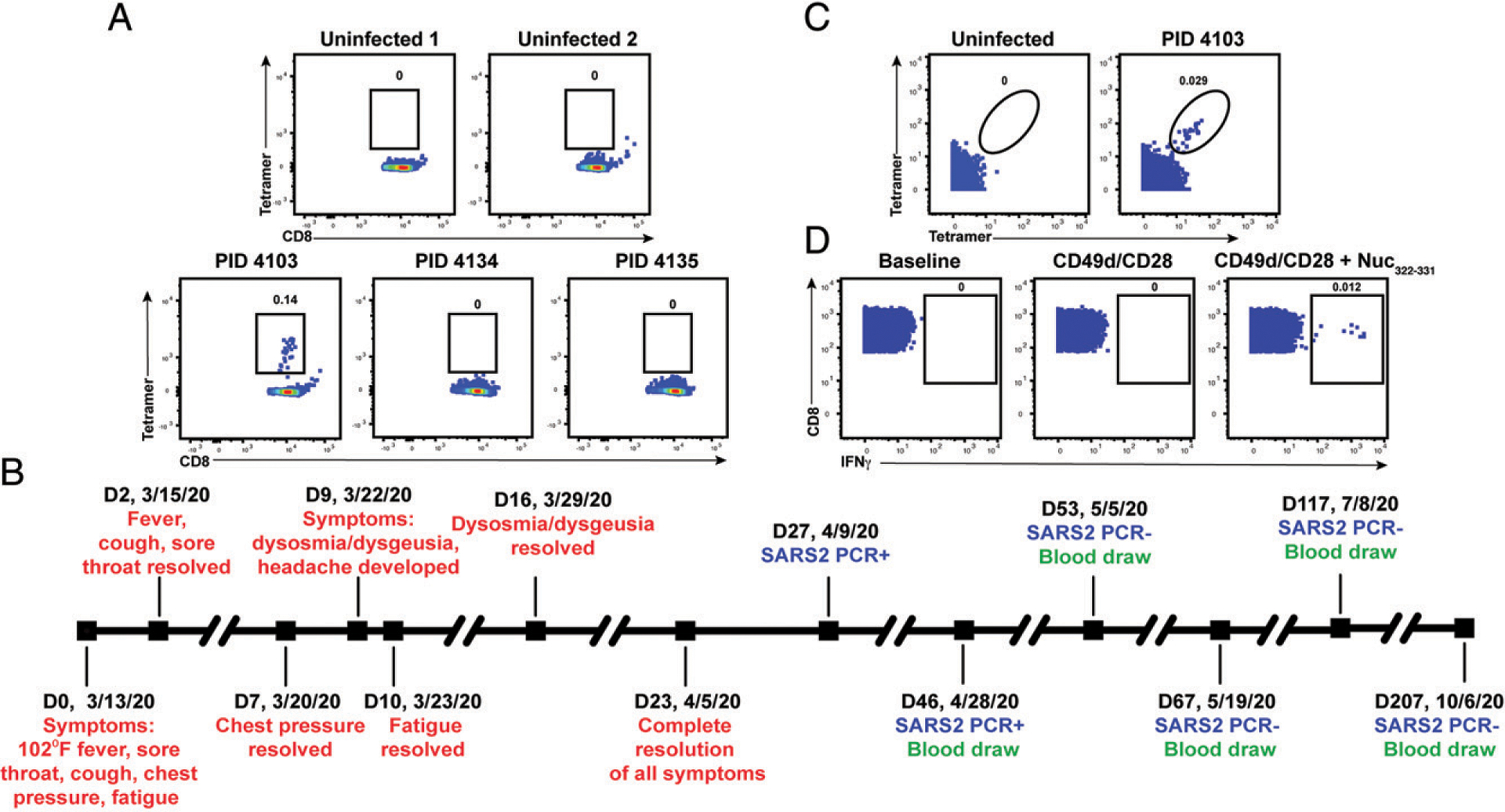
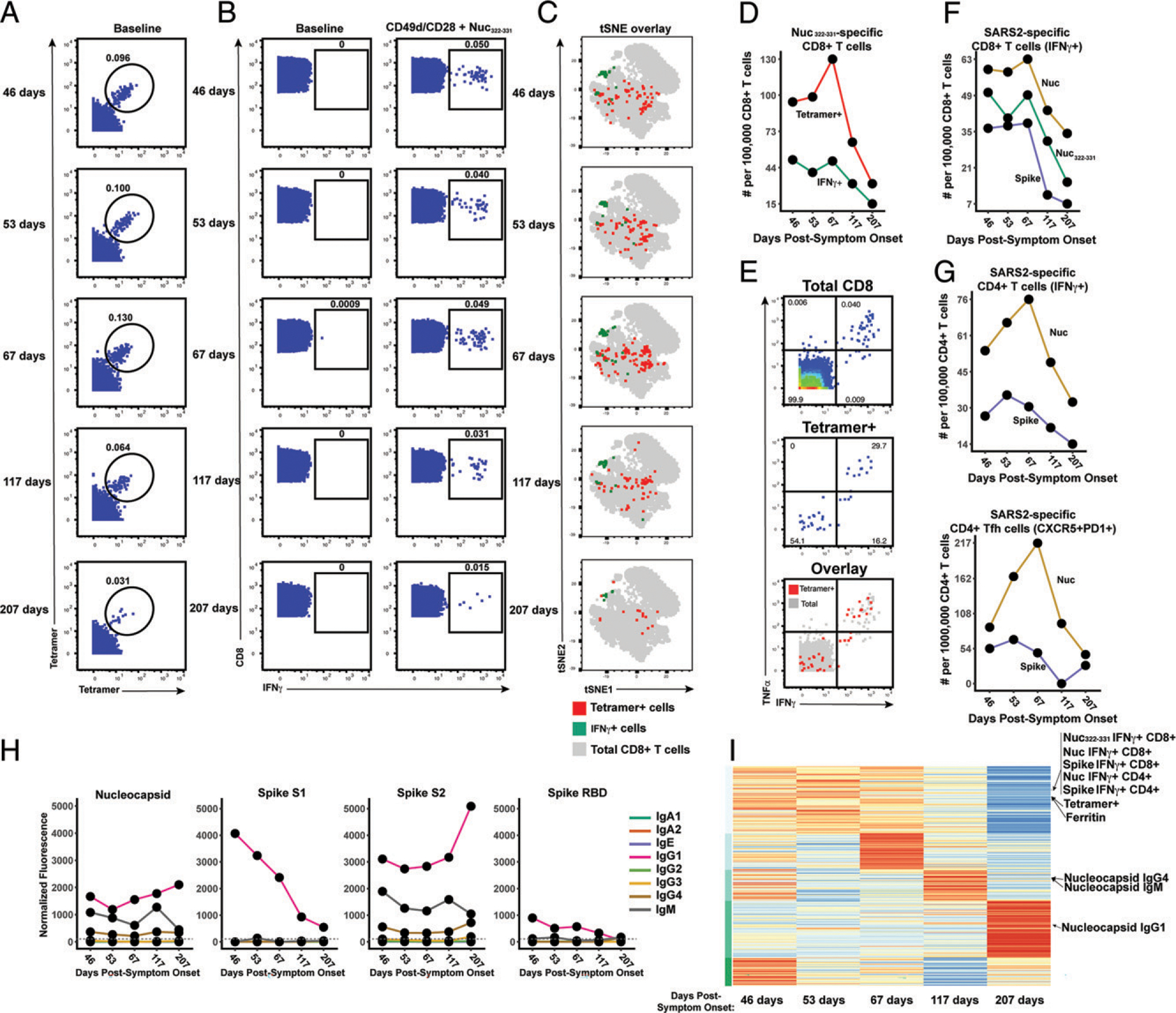
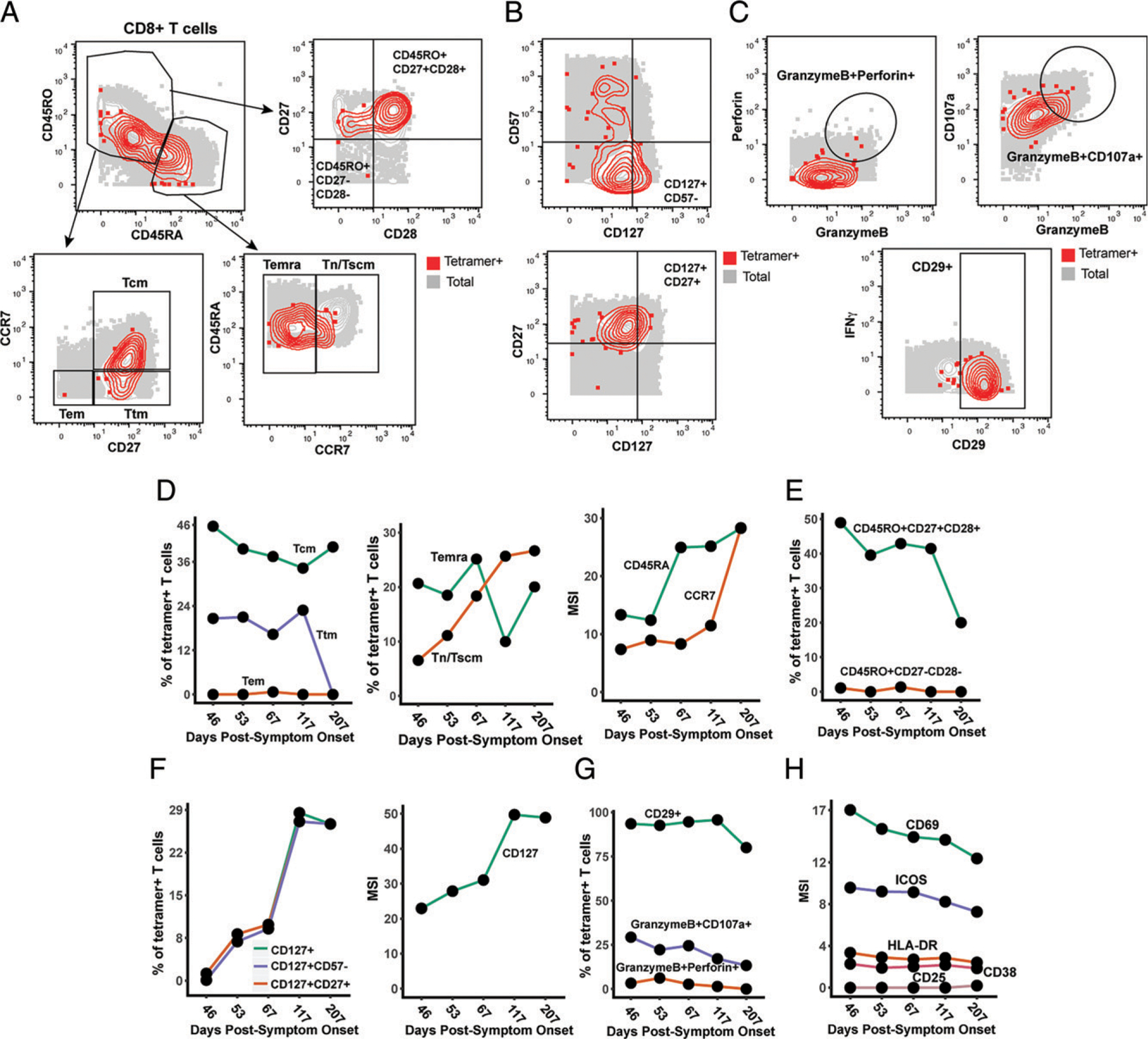
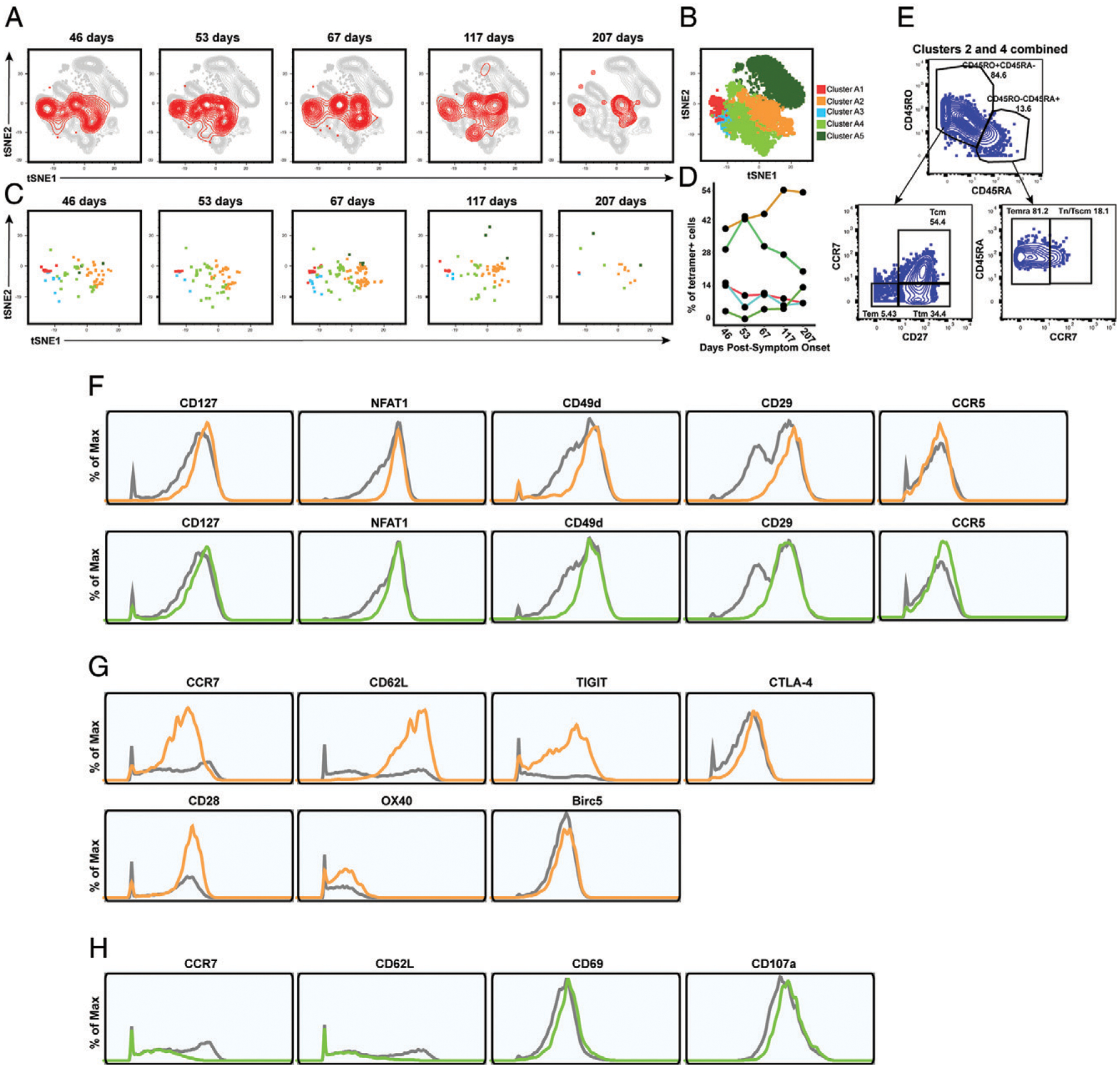
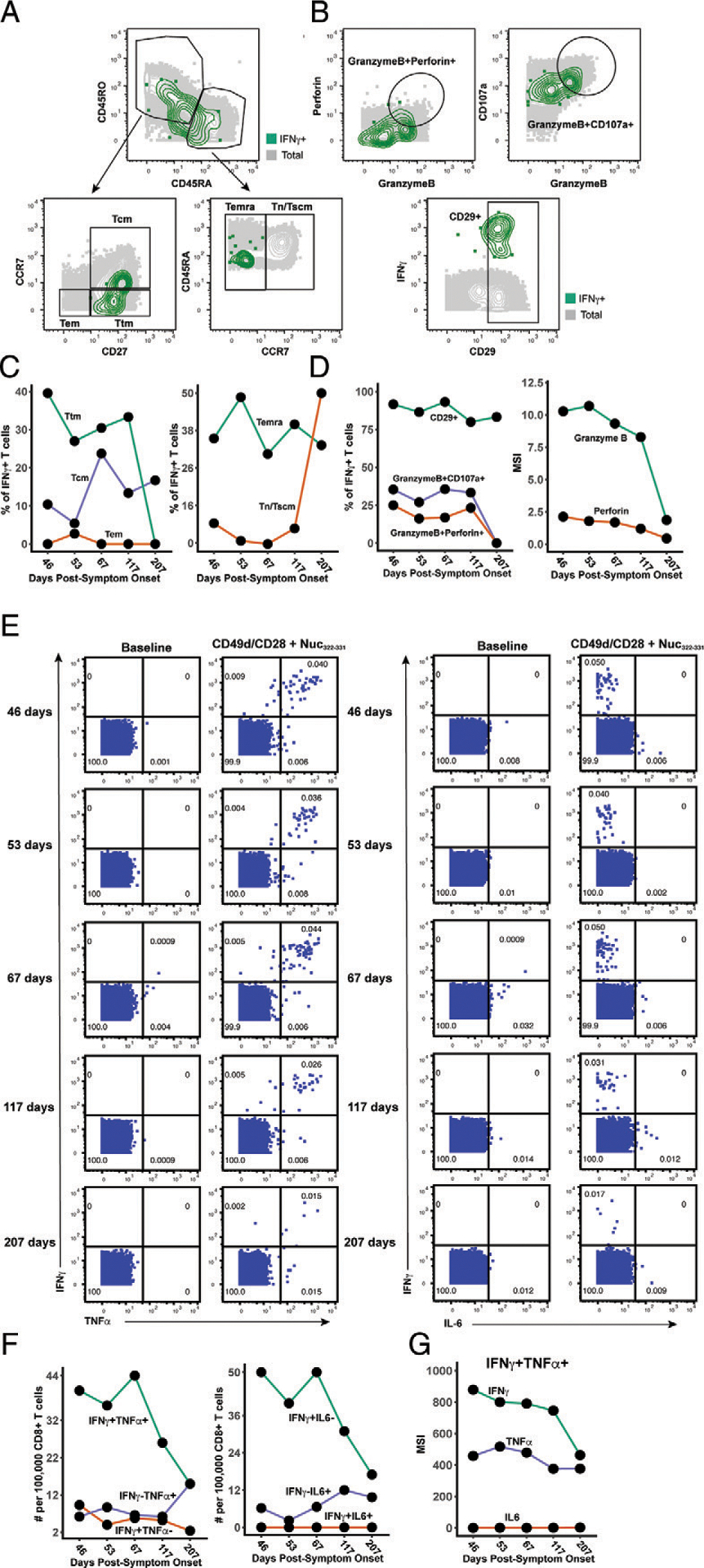
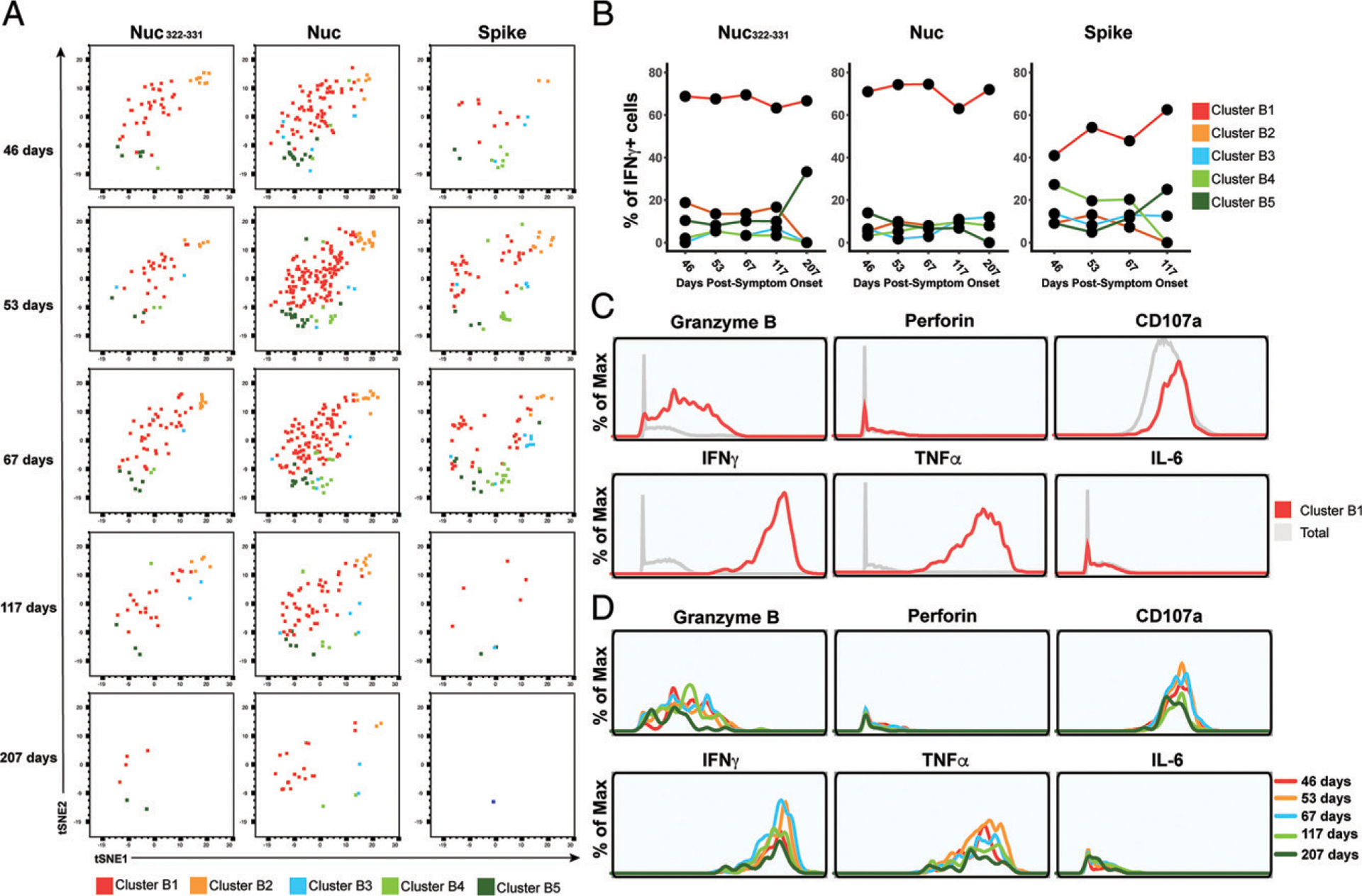
CD8+ T cells can potentiate long-lived immunity against COVID-19. We screened longitudinally-sampled convalescent human donors against SARS-CoV-2 tetramers and identified a participant with an immunodominant response against residues 322 to 311 of nucleocapsid (Nuc322-331), a peptide conserved in all variants of concern reported to date. We conducted 38-parameter cytometry by time of flight on tetramer-identified Nuc322-331-specific CD8+ T cells and on CD4+ and CD8+ T cells recognizing the entire nucleocapsid and spike proteins, and took 32 serological measurements. We discovered a coordination of the Nuc322-331-specific CD8+ T response with both the CD4+ T cell and Ab pillars of adaptive immunity. Over the approximately six month period of convalescence monitored, we observed a slow and progressive decrease in the activation state and polyfunctionality of Nuc322-331-specific CD8+ T cells, accompanied by an increase in their lymph node-homing and homeostatic proliferation potential. These results suggest that following a typical case of mild COVID-19, SARS-CoV-2-specific CD8+ T cells not only persist but continuously differentiate in a coordinated fashion well into convalescence into a state characteristic of long-lived, self-renewing memory.
Copyright © 2021 by The American Association of Immunologists, Inc.
Conflict of interest statement
Disclosures
The authors have no financial conflicts of interest.
Figures
Update of
-
Protracted yet coordinated differentiation of long-lived SARS-CoV-2-specific CD8+ T cells during COVID-19 convalescence.bioRxiv [Preprint]. 2021 Apr 29:2021.04.28.441880. doi: 10.1101/2021.04.28.441880. bioRxiv. 2021. Update in: J Immunol. 2021 Sep 1;207(5):1344-1356. doi: 10.4049/jimmunol.2100465. PMID: 33948597 Free PMC article. Updated. Preprint.
References
Publication types
MeSH terms
Grants and funding
- 75N92020D00005/HL/NHLBI NIH HHS/United States
- S10 1S10OD018040-01/OD/NIH HHS/United States
- P30 DK063720/DK/NIDDK NIH HHS/United States
- R01 AI123126/AI/NIAID NIH HHS/United States
- 75N93020C00005/AI/NIAID NIH HHS/United States
- 75N93023D00005/AI/NIAID NIH HHS/United States
- 75N95020D00005/DA/NIDA NIH HHS/United States
- P30 AI027763/AI/NIAID NIH HHS/United States
- 75N99020D00005/OF/ORFDO NIH HHS/United States
- S10 OD018040/OD/NIH HHS/United States
- R01 AI121207/AI/NIAID NIH HHS/United States
- T32 AI007334/AI/NIAID NIH HHS/United States
- 75N93022D00005/AI/NIAID NIH HHS/United States
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
