

Spinal lumbar dI2 interneurons contribute to stability of bipedal stepping
- PMID: 34396953
- PMCID: PMC8448531
- DOI: 10.7554/eLife.62001
Spinal lumbar dI2 interneurons contribute to stability of bipedal stepping
Abstract
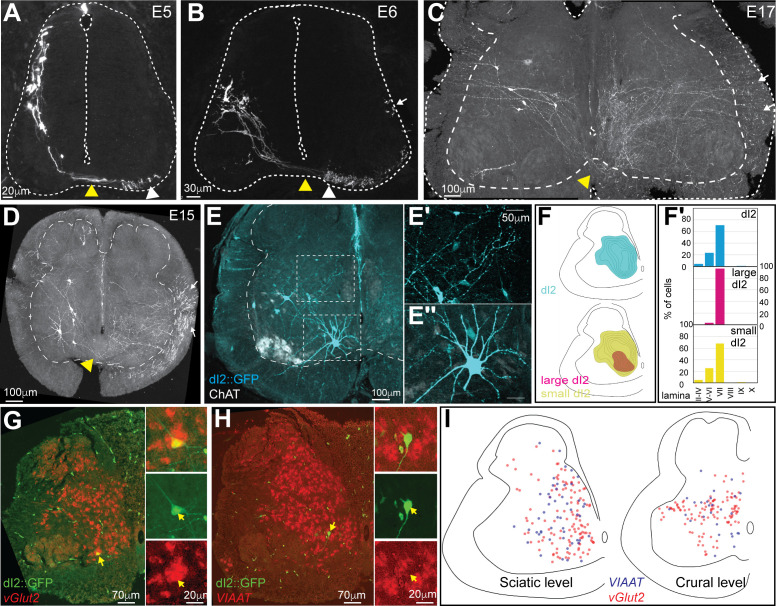
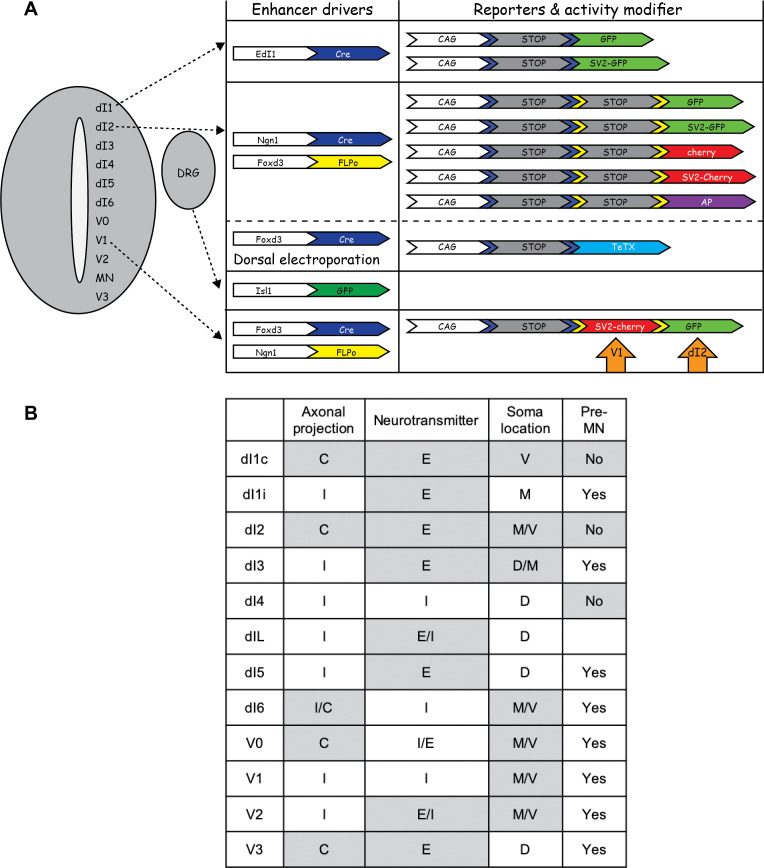
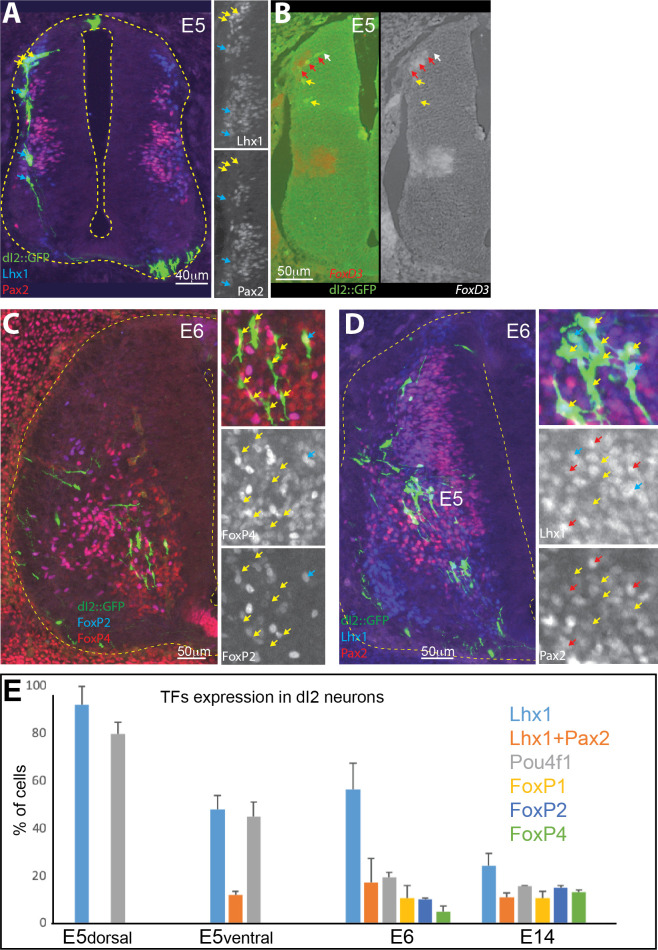
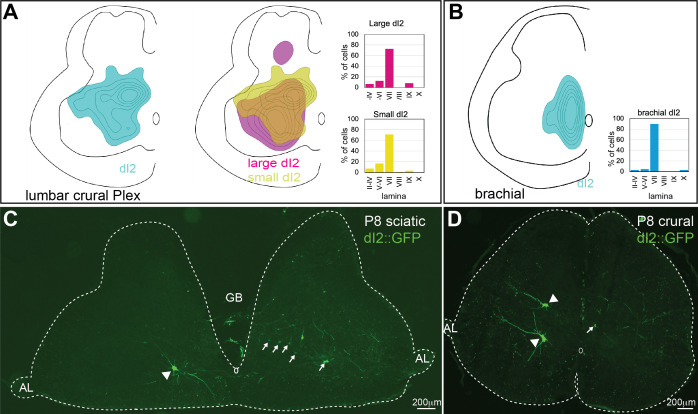
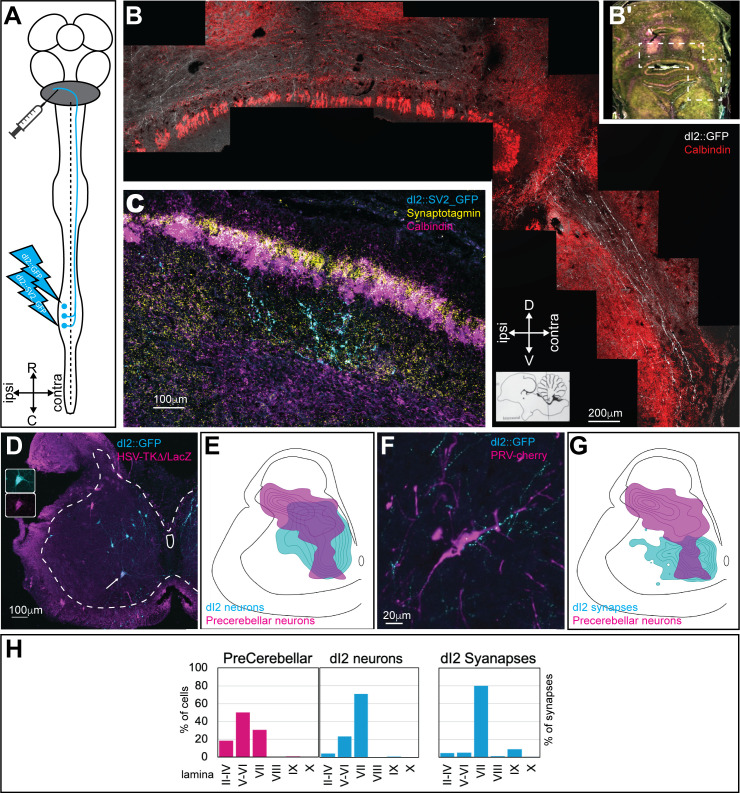
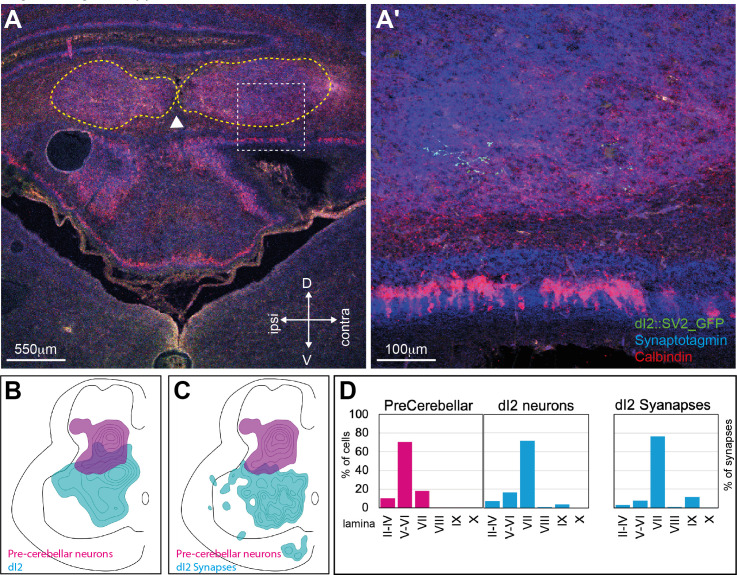
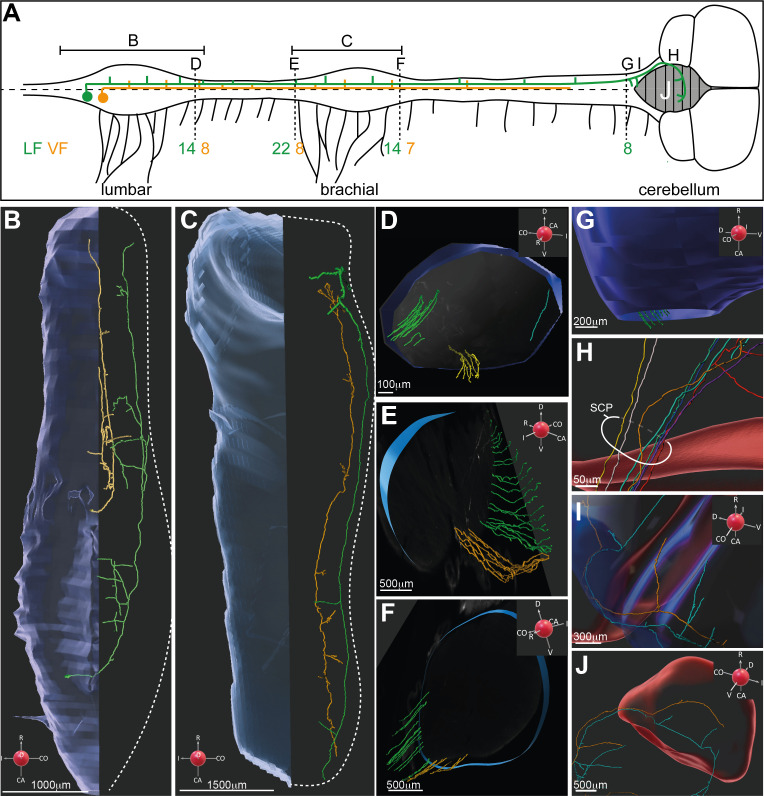
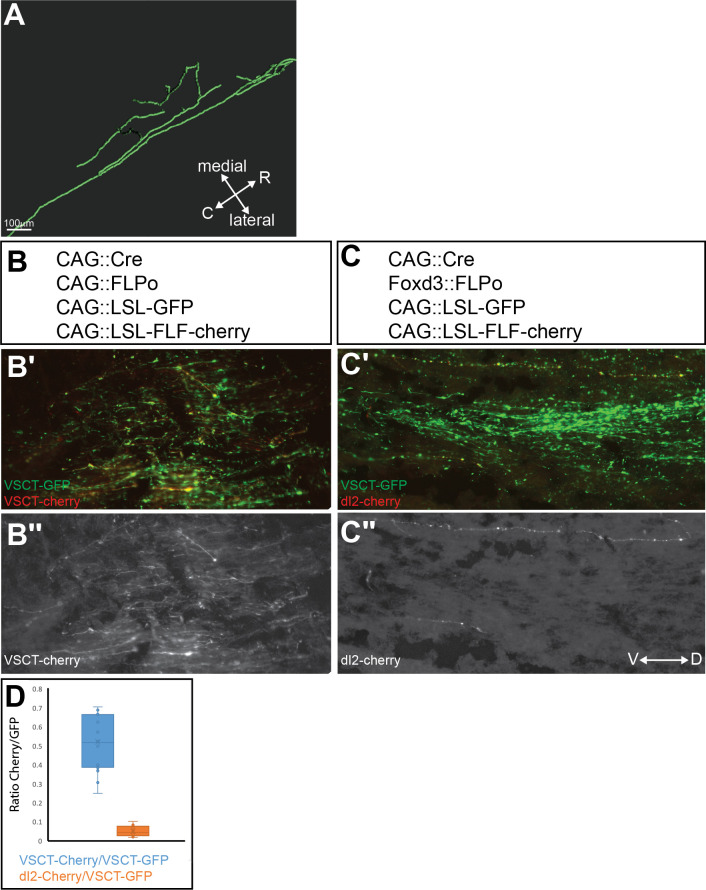
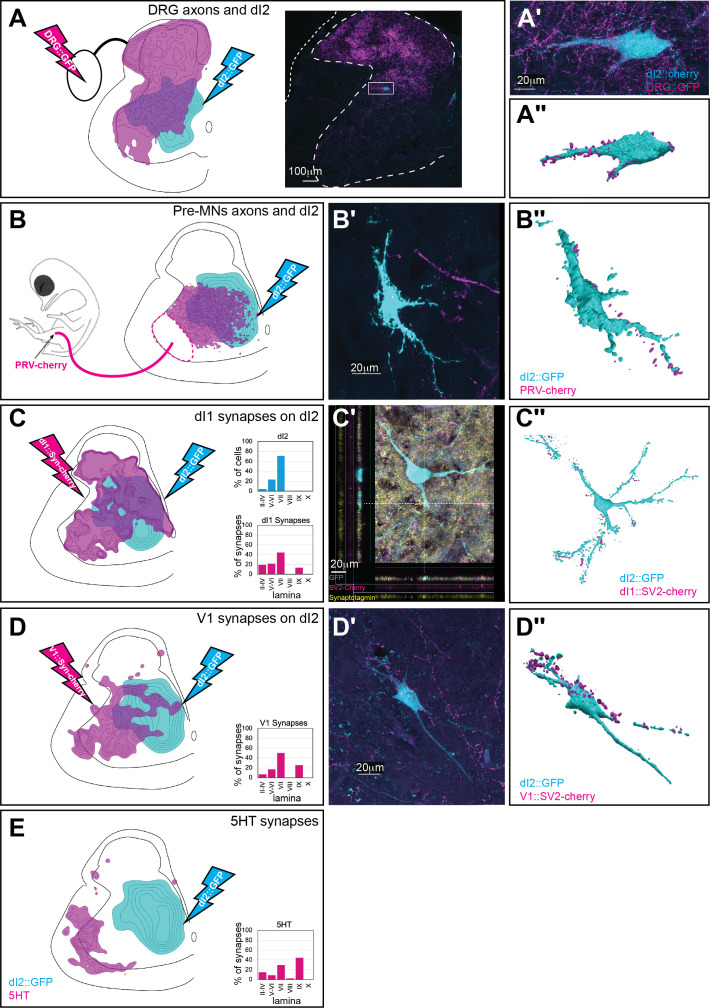
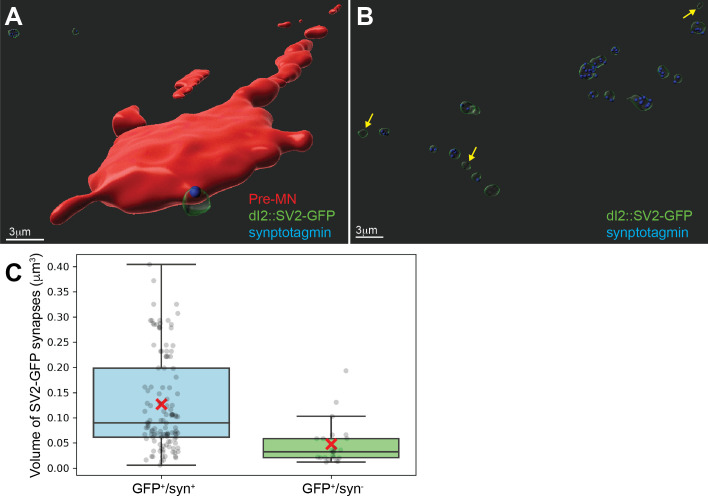
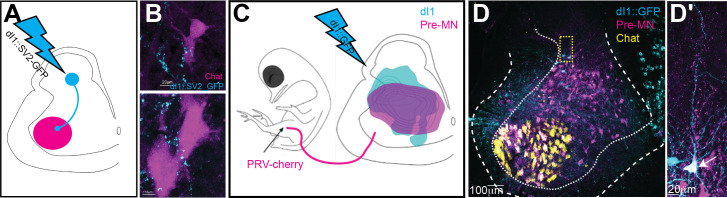
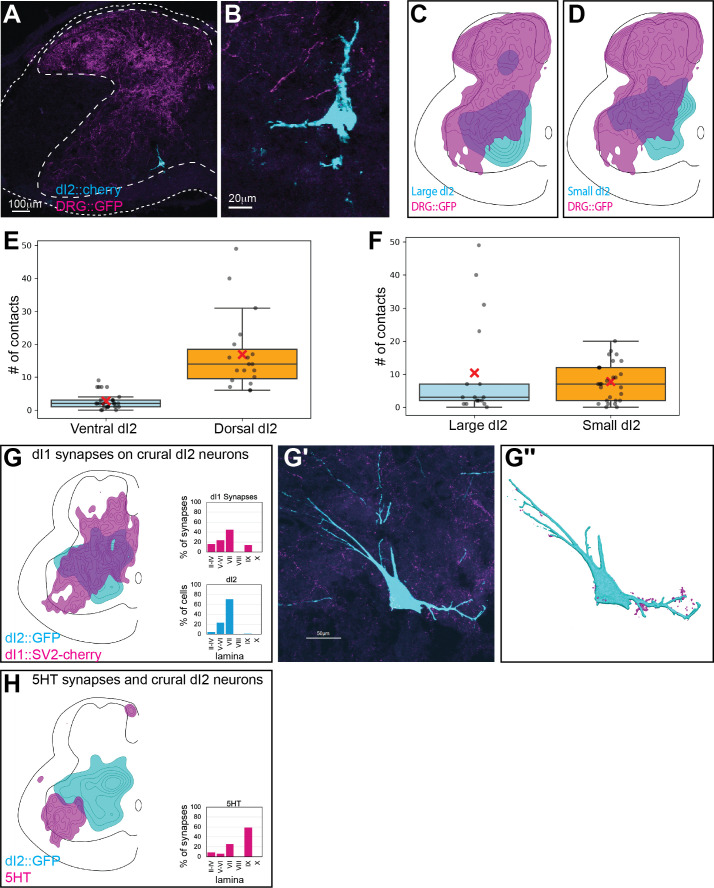
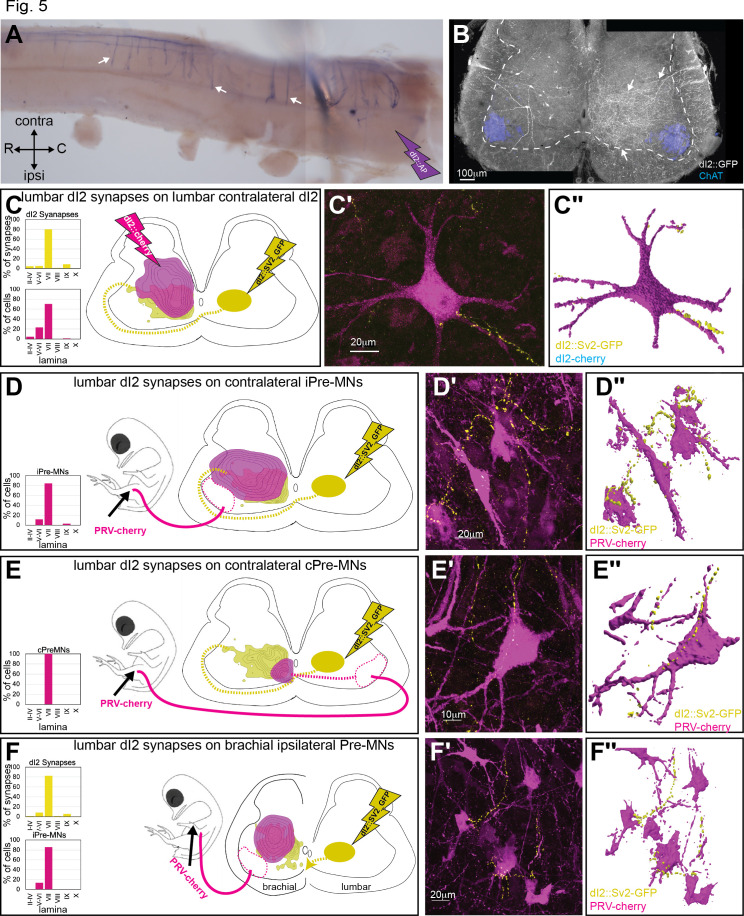
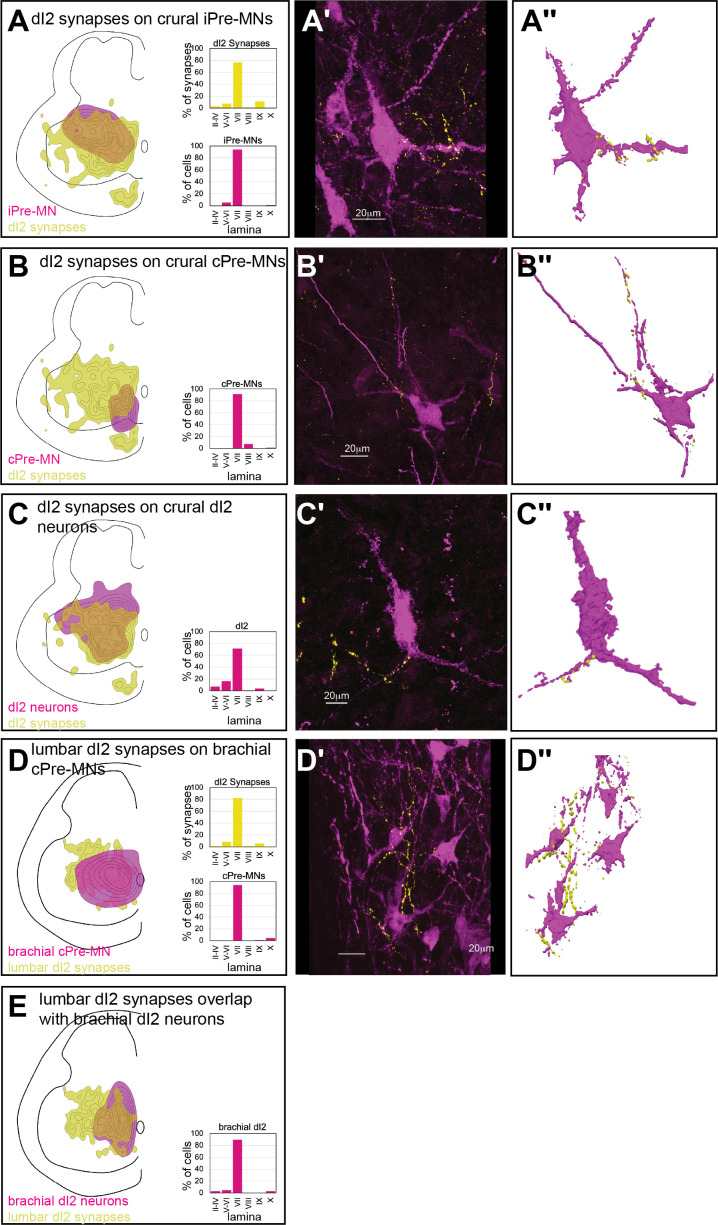
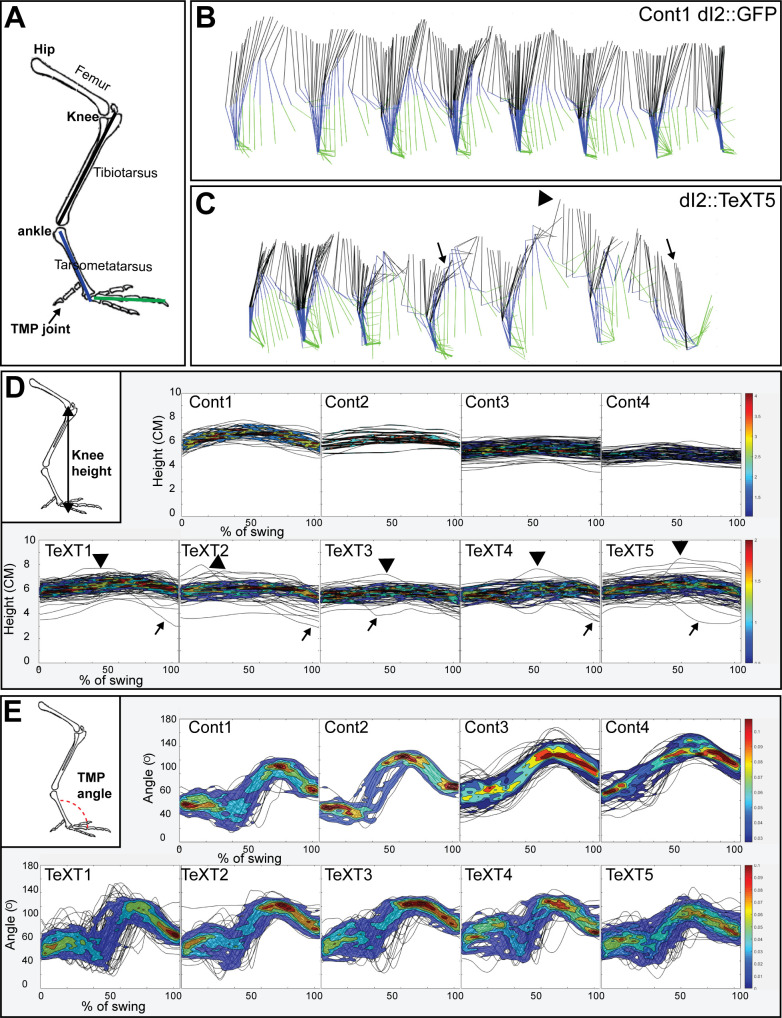
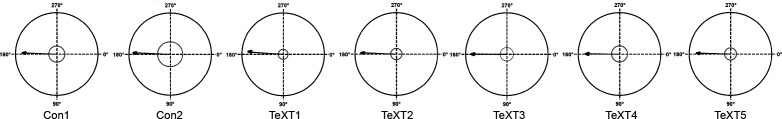
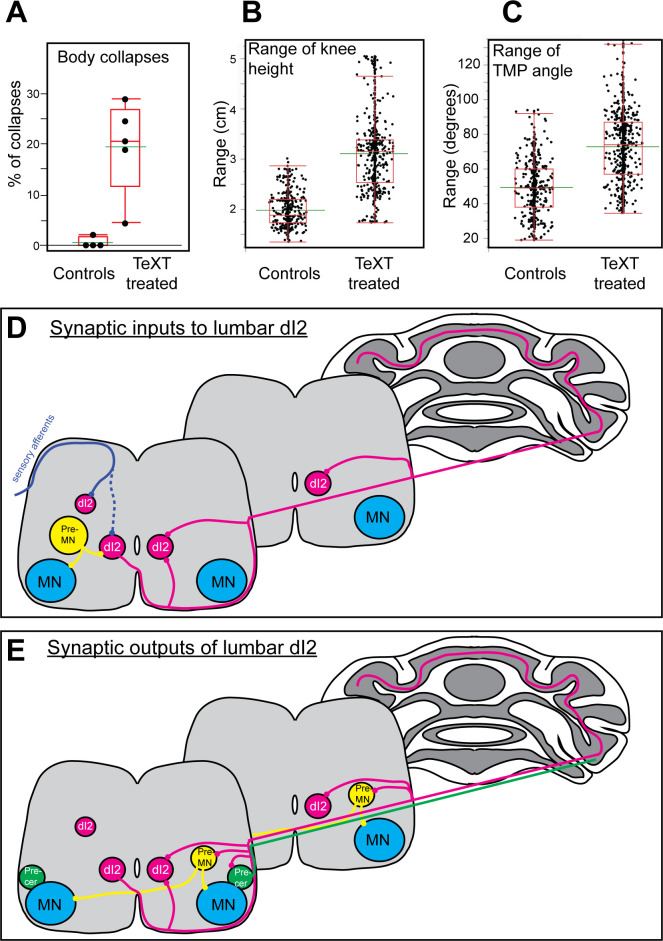
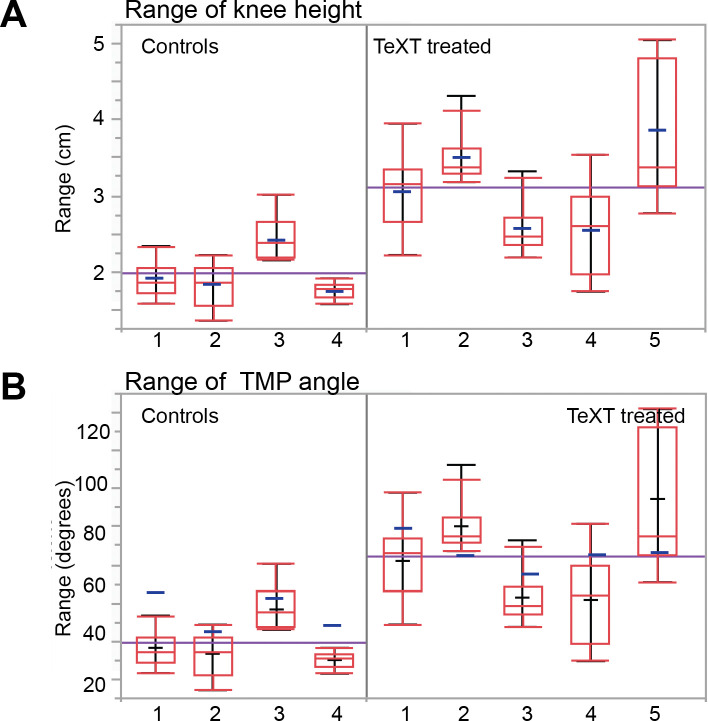
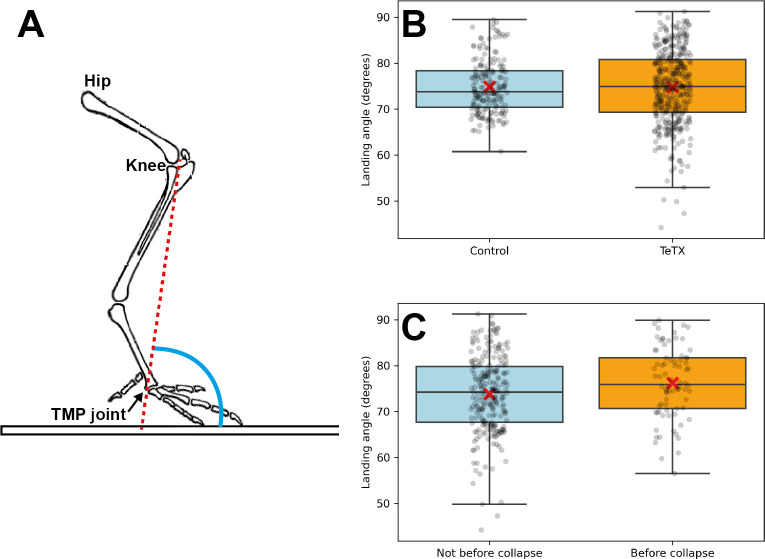
Peripheral and intraspinal feedback is required to shape and update the output of spinal networks that execute motor behavior. We report that lumbar dI2 spinal interneurons in chicks receive synaptic input from afferents and premotor neurons. These interneurons innervate contralateral premotor networks in the lumbar and brachial spinal cord, and their ascending projections innervate the cerebellum. These findings suggest that dI2 neurons function as interneurons in local lumbar circuits, are involved in lumbo-brachial coupling, and that part of them deliver peripheral and intraspinal feedback to the cerebellum. Silencing of dI2 neurons leads to destabilized stepping in posthatching day 8 hatchlings, with occasional collapses, variable step profiles, and a wide-base walking gait, suggesting that dI2 neurons may contribute to the stabilization of the bipedal gait.
Keywords: cerebellum; chicken; developmental biology; interneurons; locomotion; neural circuits; neuroscience; spinal cord; spinocerebellar tract.
© 2021, Haimson et al.
Conflict of interest statement
BH, YH, NB, AK, MD, YC, AL, AK none
Figures
References
-
- Andersson LS, Larhammar M, Memic F, Wootz H, Schwochow D, Rubin CJ, Patra K, Arnason T, Wellbring L, Hjälm G, Imsland F, Petersen JL, McCue ME, Mickelson JR, Cothran G, Ahituv N, Roepstorff L, Mikko S, Vallstedt A, Lindgren G, Andersson L, Kullander K. Mutations in DMRT3 affect locomotion in horses and spinal circuit function in mice. Nature. 2012;488:642–646. doi: 10.1038/nature11399. - DOI - PMC - PubMed
