

A therapeutic cancer vaccine delivers antigens and adjuvants to lymphoid tissues using genetically modified T cells
- PMID: 34396986
- PMCID: PMC8363286
- DOI: 10.1172/JCI144195
A therapeutic cancer vaccine delivers antigens and adjuvants to lymphoid tissues using genetically modified T cells
Abstract
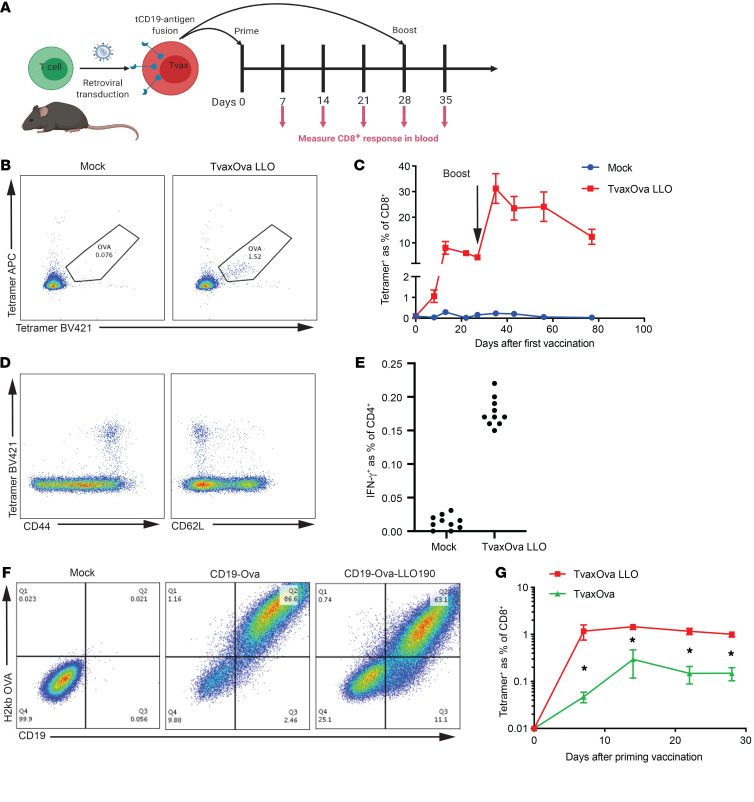
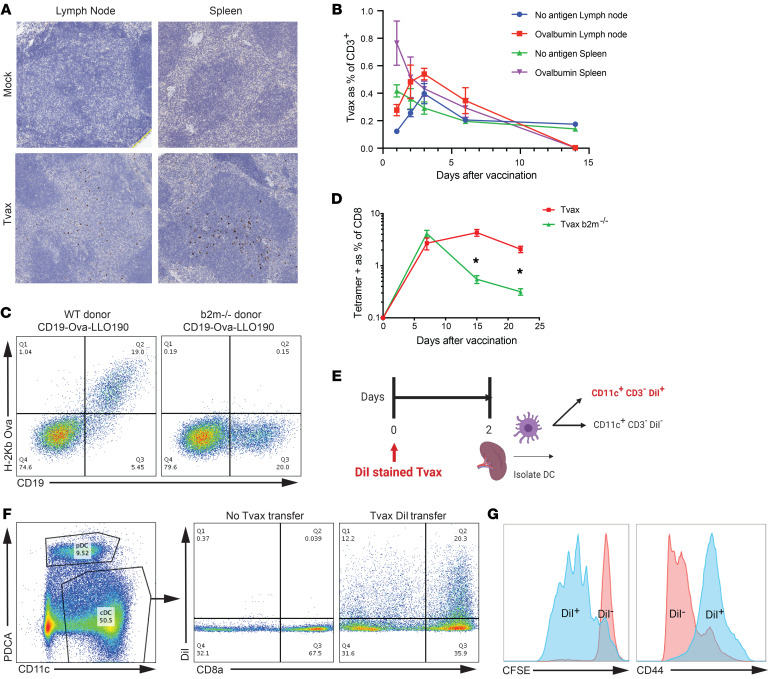
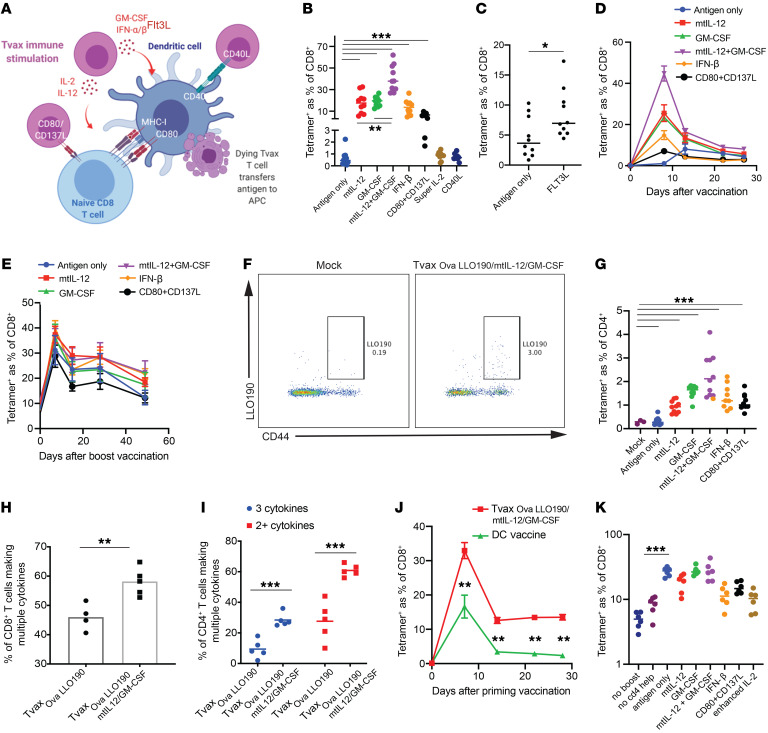
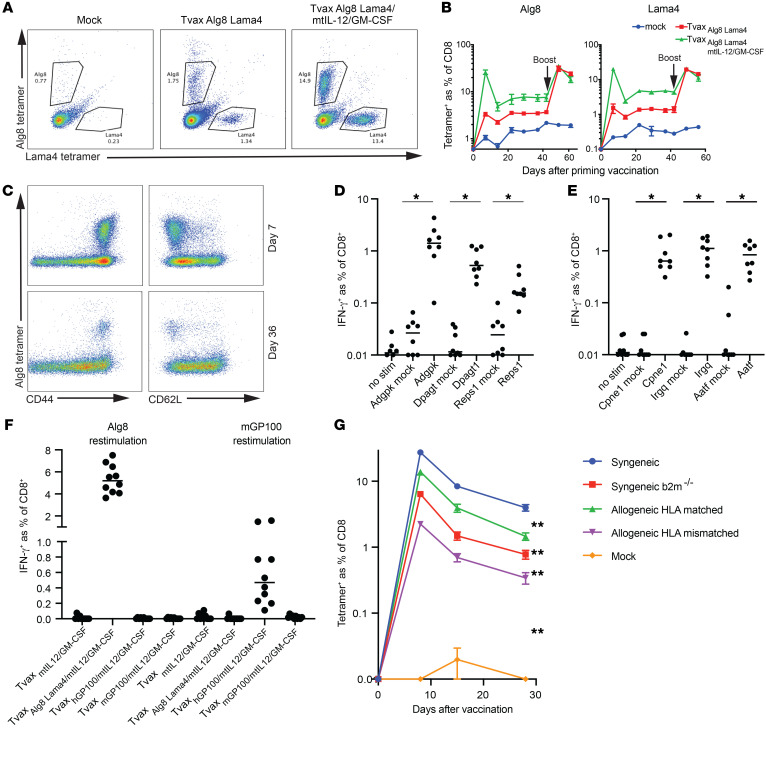
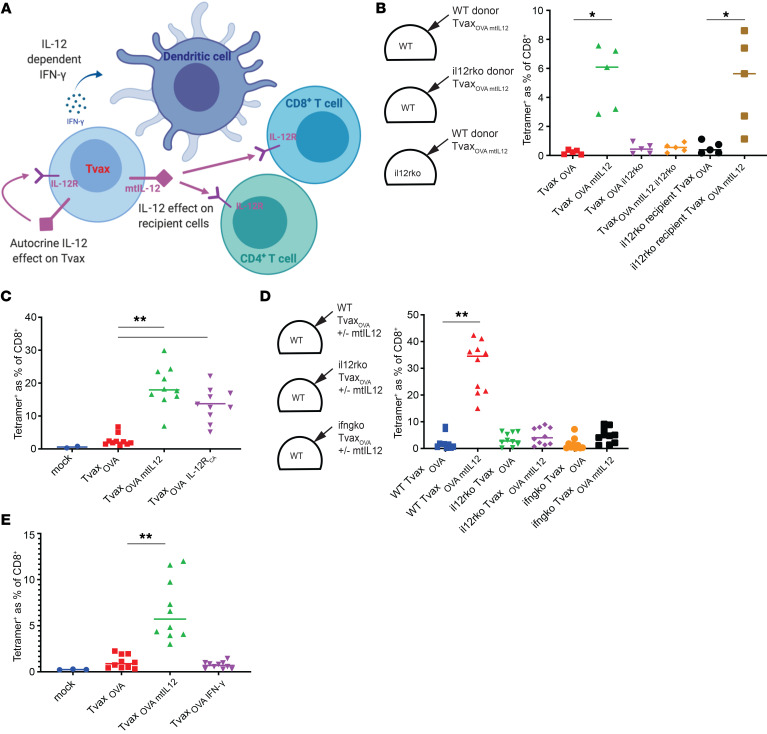
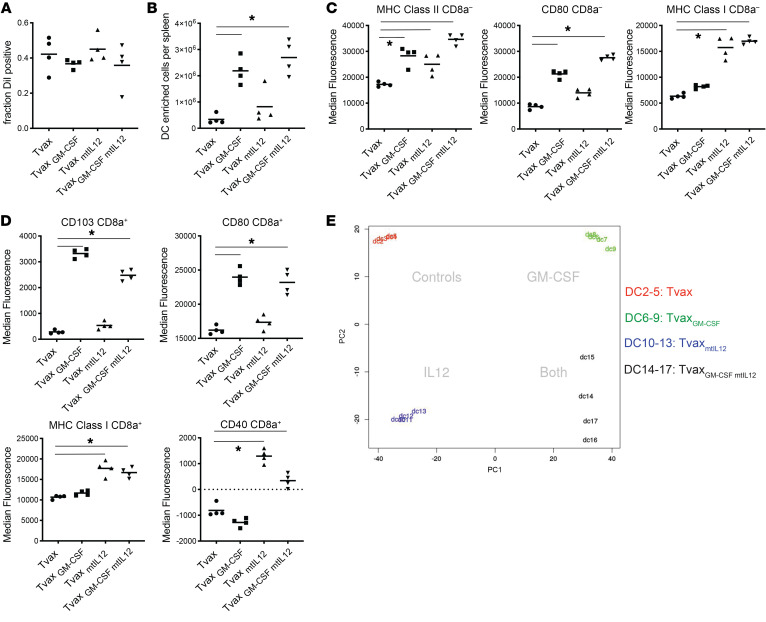
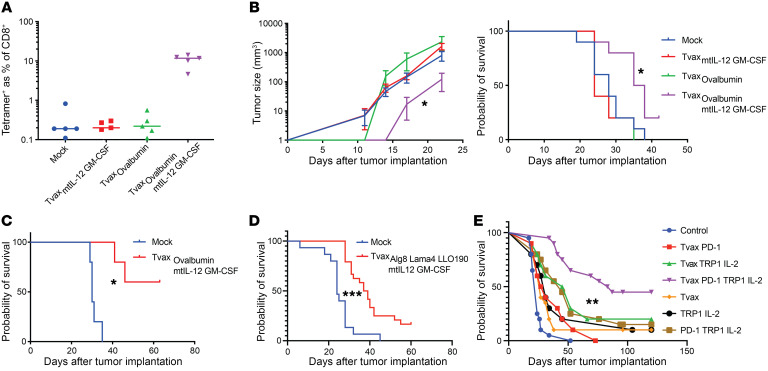
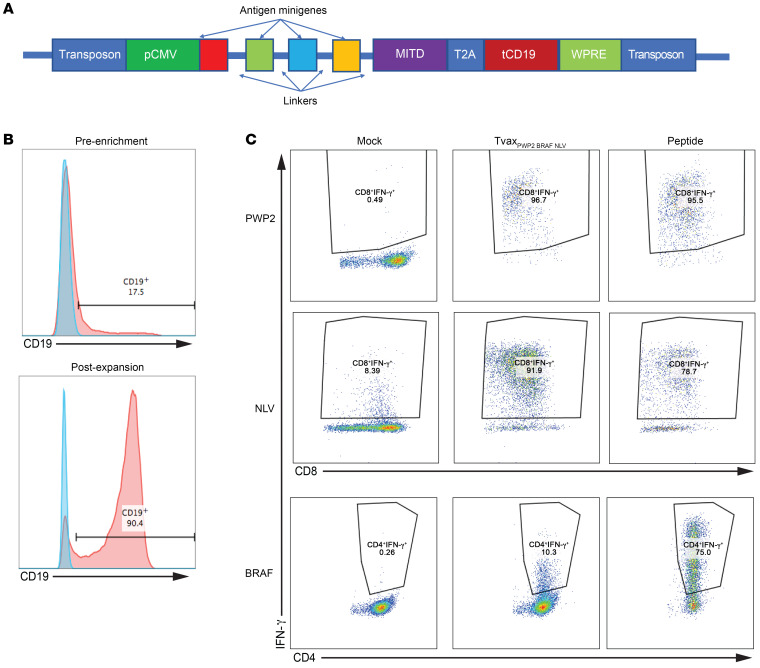
Therapeutic vaccines that augment T cell responses to tumor antigens have been limited by poor potency in clinical trials. In contrast, the transfer of T cells modified with foreign transgenes frequently induces potent endogenous T cell responses to epitopes in the transgene product, and these responses are undesirable, because they lead to rejection of the transferred T cells. We sought to harness gene-modified T cells as a vaccine platform and developed cancer vaccines composed of autologous T cells modified with tumor antigens and additional adjuvant signals (Tvax). T cells expressing model antigens and a broad range of tumor neoantigens induced robust and durable T cell responses through cross-presentation of antigens by host DCs. Providing Tvax with signals such as CD80, CD137L, IFN-β, IL-12, GM-CSF, and FLT3L enhanced T cell priming. Coexpression of IL-12 and GM-CSF induced the strongest CD4+ and CD8+ T cell responses through complimentary effects on the recruitment and activation of DCs, mediated by autocrine IL-12 receptor signaling in the Tvax. Therapeutic vaccination with Tvax and adjuvants showed antitumor activity in subcutaneous and metastatic preclinical mouse models. Human T cells modified with neoantigens readily activated specific T cells derived from patients, providing a path for clinical translation of this therapeutic platform in cancer.
Keywords: Cancer immunotherapy; Immunology; Vaccines.
Conflict of interest statement
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
