

Generation of Naïve Blastoderm Explants from Zebrafish Embryos
- PMID: 34398141
- PMCID: PMC8892589
- DOI: 10.3791/62797
Generation of Naïve Blastoderm Explants from Zebrafish Embryos
Abstract
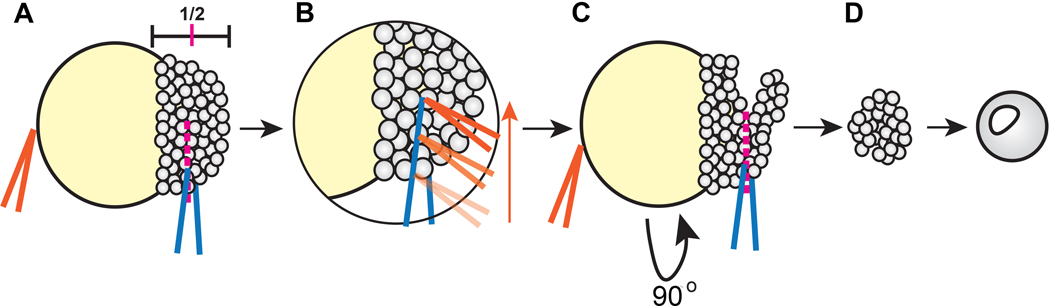
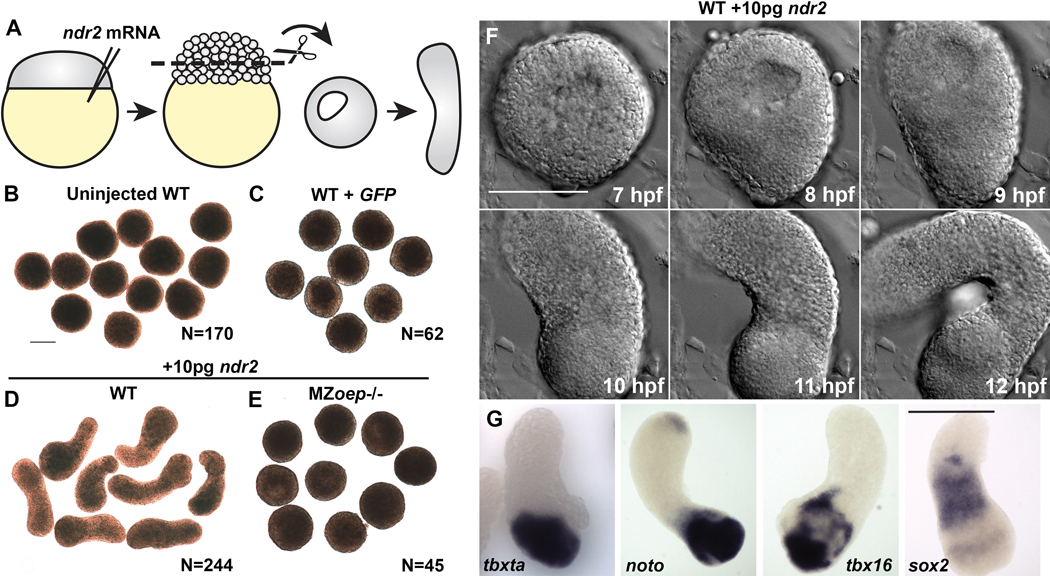
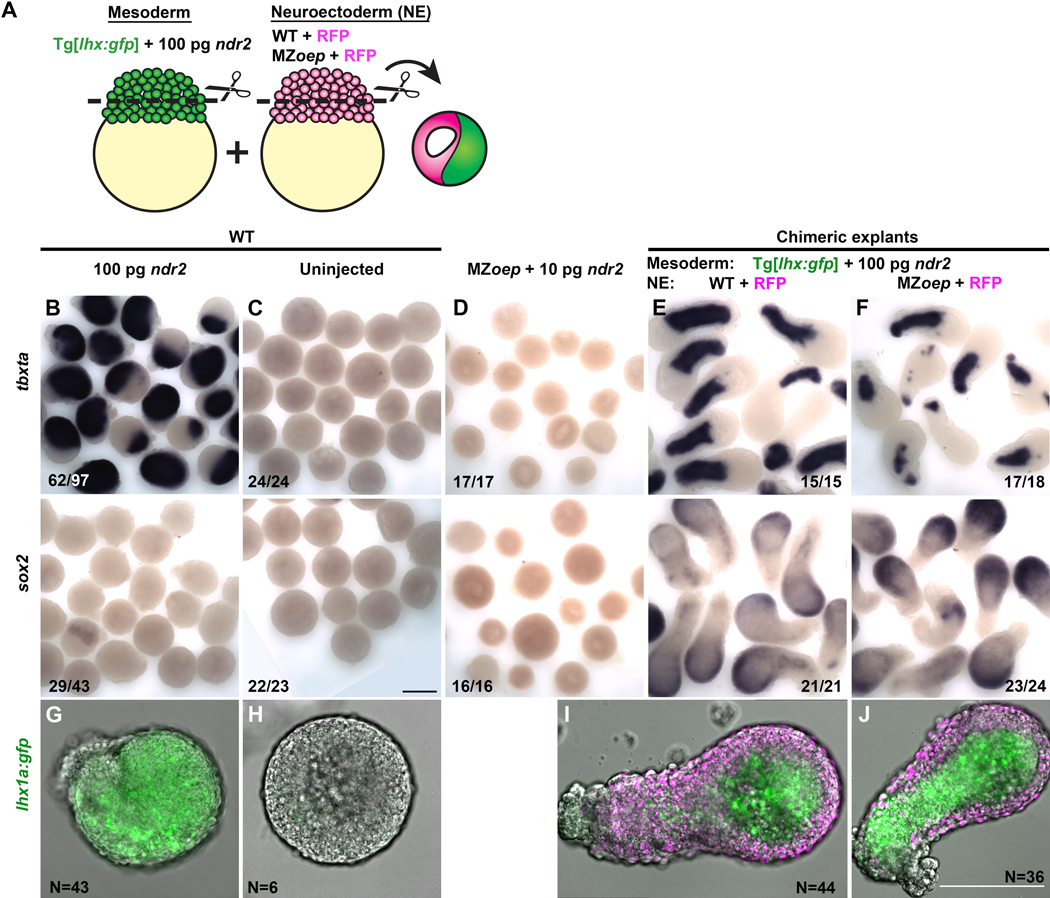
Due to their optical clarity and rapid development, zebrafish embryos are an excellent system for examining cell behaviors and developmental processes. However, because of the complexity and redundancy of embryonic signals, it can be challenging to discern the complete role of any single signal during early embryogenesis. By explanting the animal region of the zebrafish blastoderm, relatively naïve clusters of embryonic cells are generated that can be easily cultured and manipulated ex vivo. By introducing a gene of interest by RNA injection before explantation, one can assess the effect of this molecule on gene expression, cell behaviors, and other developmental processes in relative isolation. Furthermore, cells from embryos of different genotypes or conditions can be combined in a single chimeric explant to examine cell/tissue interactions and tissue-specific gene functions. This article provides instructions for generating zebrafish blastoderm explants and demonstrates that a single signaling molecule - a Nodal ligand - is sufficient to induce germ layer formation and extension morphogenesis in otherwise naïve embryonic tissues. Due to their ability to recapitulate embryonic cell behaviors, morphogen gradients, and gene expression patterns in a simplified ex vivo system, these explants are anticipated to be of great utility to many zebrafish researchers.
Figures
References
-
- Green JB, New HV, Smith JC Responses of embryonic Xenopus cells to activin and FGF are separated by multiple dose thresholds and correspond to distinct axes of the mesoderm. Cell. 71 (5), 731–739 (1992). - PubMed
-
- Sudarwati S, Nieuwkoop PD Mesoderm formation in the anuranXenopus laevis (Daudin). Wilhelm Roux Archiv fur Entwicklungsmechanik der Organismen. 166 (3), 189–204 (1971). - PubMed
-
- Gurdon JB, Fairman S, Mohun TJ, Brennan S. Activation of muscle-specific actin genes in Xenopus development by an induction between animal and vegetal cells of a blastula. Cell. 41 (3), 913–922 (1985). - PubMed
-
- Keller R, Danilchik M. Regional expression, pattern and timing of convergence and extension during gastrulation of Xenopus laevis. Development. 103 (1), 193–209 (1988). - PubMed
-
- Ariizumi T. et al. Isolation and differentiation of Xenopus animal cap cells. Current Protocols in Stem Cell Biology. Chapter 1 Unit 1D.5 (2009). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
