

Fourteen Stations of Auxin
- PMID: 34400554
- PMCID: PMC9159264
- DOI: 10.1101/cshperspect.a039859
Fourteen Stations of Auxin
Abstract
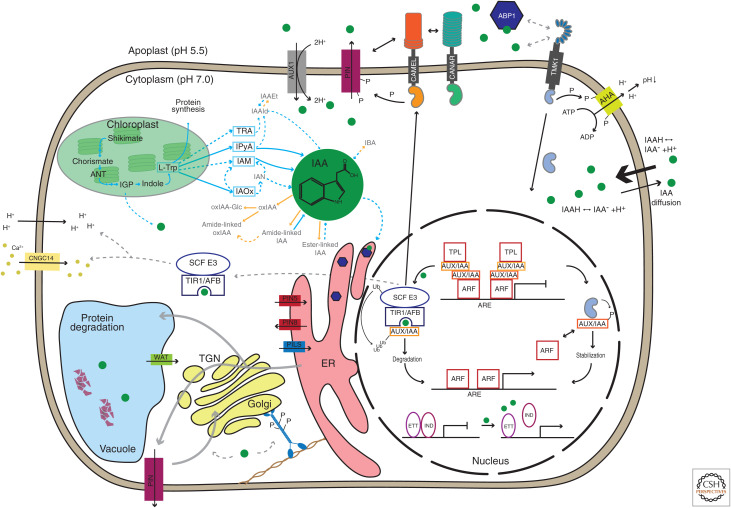
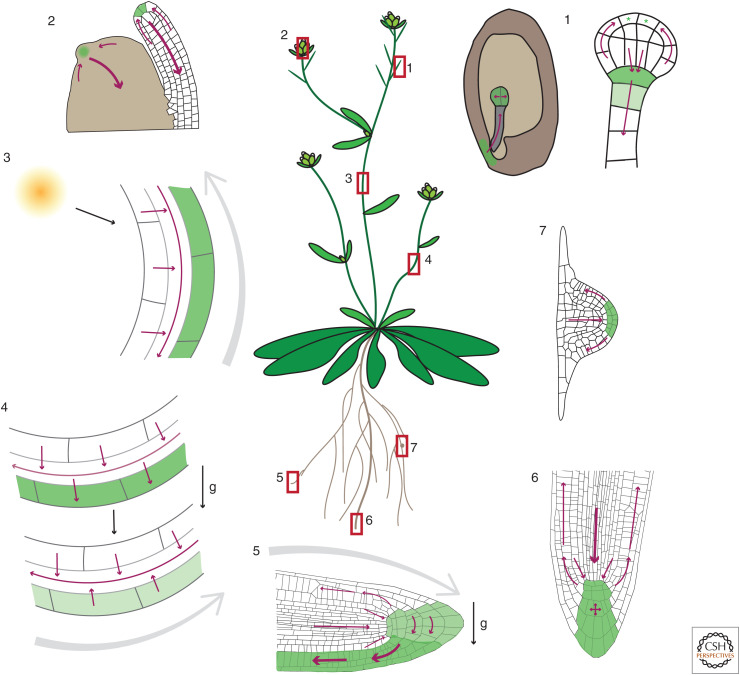
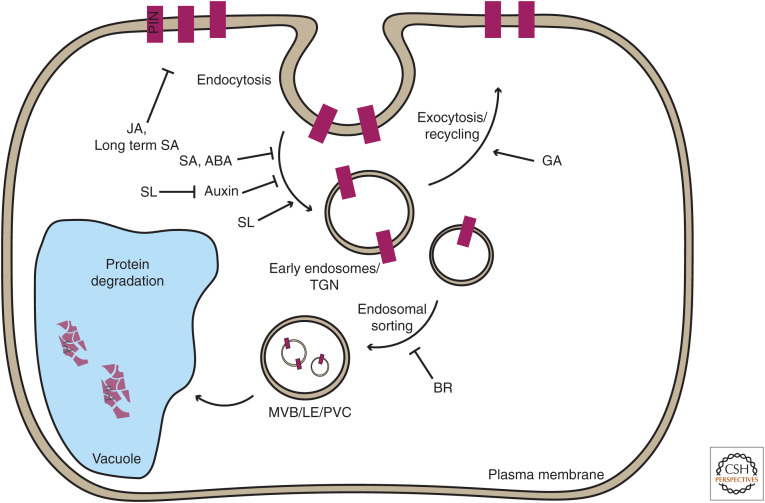
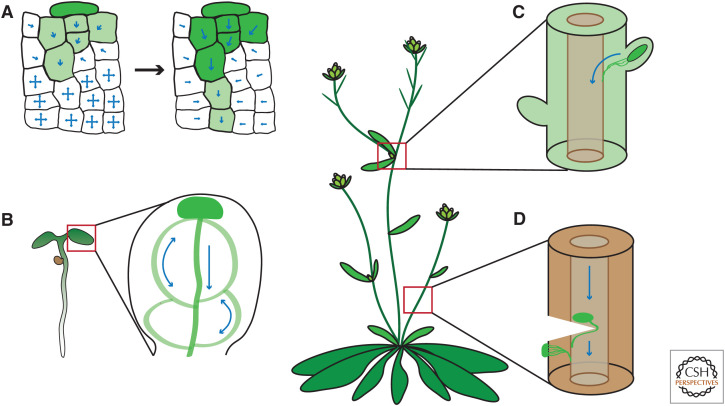
Auxin has always been at the forefront of research in plant physiology and development. Since the earliest contemplations by Julius von Sachs and Charles Darwin, more than a century-long struggle has been waged to understand its function. This largely reflects the failures, successes, and inevitable progress in the entire field of plant signaling and development. Here I present 14 stations on our long and sometimes mystical journey to understand auxin. These highlights were selected to give a flavor of the field and to show the scope and limits of our current knowledge. A special focus is put on features that make auxin unique among phytohormones, such as its dynamic, directional transport network, which integrates external and internal signals, including self-organizing feedback. Accented are persistent mysteries and controversies. The unexpected discoveries related to rapid auxin responses and growth regulation recently disturbed our contentment regarding understanding of the auxin signaling mechanism. These new revelations, along with advances in technology, usher us into a new, exciting era in auxin research.
Copyright © 2022 Cold Spring Harbor Laboratory Press; all rights reserved.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources