

Nuclear Compartments: An Incomplete Primer to Nuclear Compartments, Bodies, and Genome Organization Relative to Nuclear Architecture
- PMID: 34400557
- PMCID: PMC9248822
- DOI: 10.1101/cshperspect.a041268
Nuclear Compartments: An Incomplete Primer to Nuclear Compartments, Bodies, and Genome Organization Relative to Nuclear Architecture
Abstract
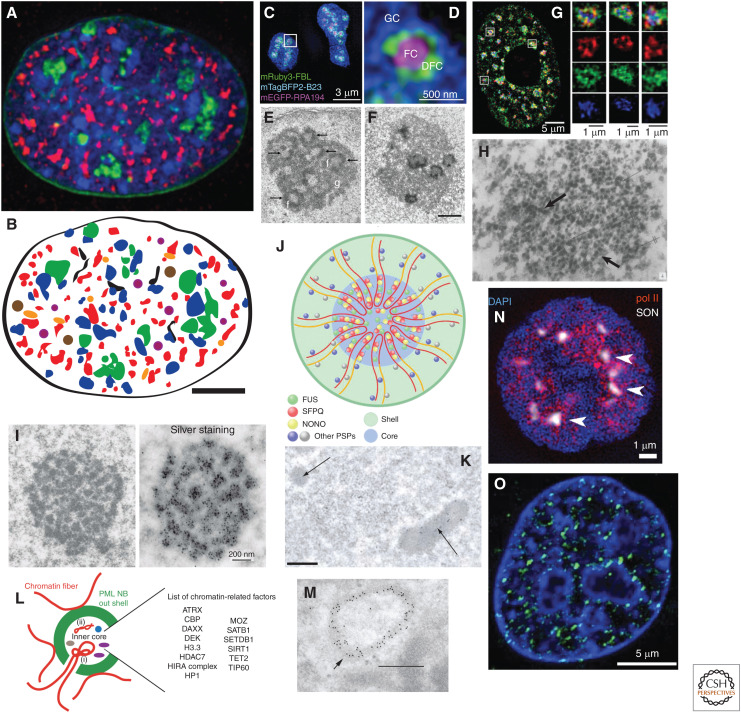
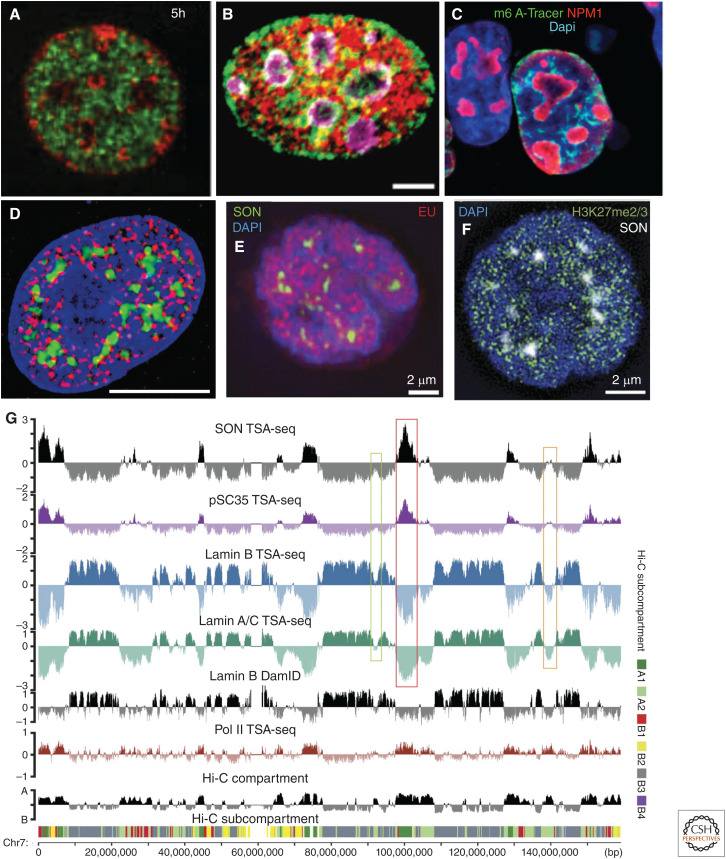
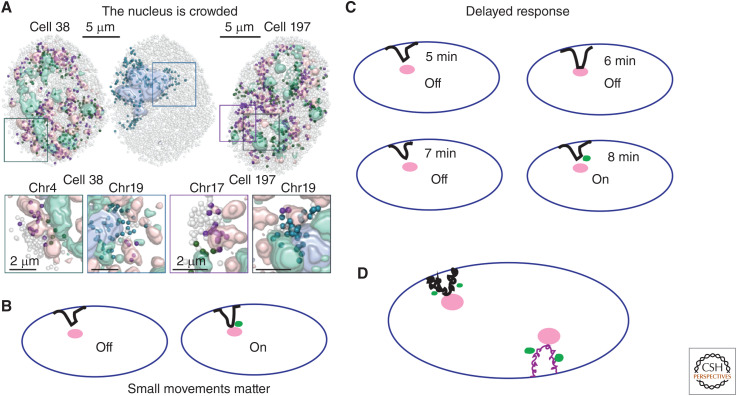
This work reviews nuclear compartments, defined broadly to include distinct nuclear structures, bodies, and chromosome domains. It first summarizes original cytological observations before comparing concepts of nuclear compartments emerging from microscopy versus genomic approaches and then introducing new multiplexed imaging approaches that promise in the future to meld both approaches. I discuss how previous models of radial distribution of chromosomes or the binary division of the genome into A and B compartments are now being refined by the recognition of more complex nuclear compartmentalization. The poorly understood question of how these nuclear compartments are established and maintained is then discussed, including through the modern perspective of phase separation, before moving on to address possible functions of nuclear compartments, using the possible role of nuclear speckles in modulating gene expression as an example. Finally, the review concludes with a discussion of future questions for this field.
Copyright © 2022 Cold Spring Harbor Laboratory Press; all rights reserved.
Figures
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources