

Coeloglossum viride var. bracteatum extract attenuates staurosporine induced neurotoxicity by restoring the FGF2-PI3K/Akt signaling axis and Dnmt3
- PMID: 34401557
- PMCID: PMC8353313
- DOI: 10.1016/j.heliyon.2021.e07503
Coeloglossum viride var. bracteatum extract attenuates staurosporine induced neurotoxicity by restoring the FGF2-PI3K/Akt signaling axis and Dnmt3
Abstract
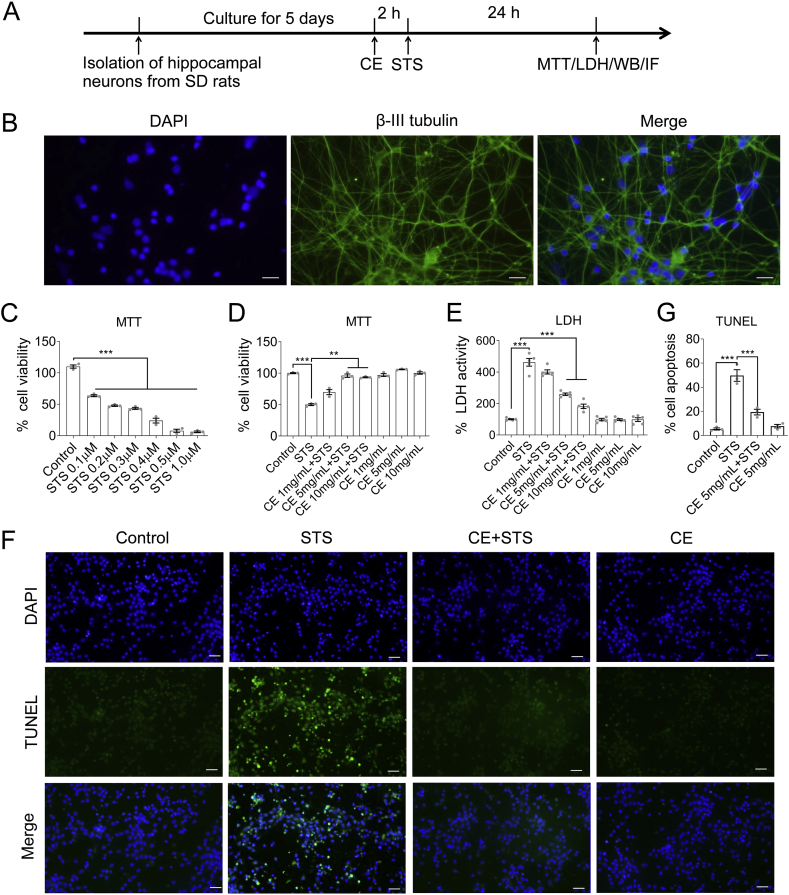
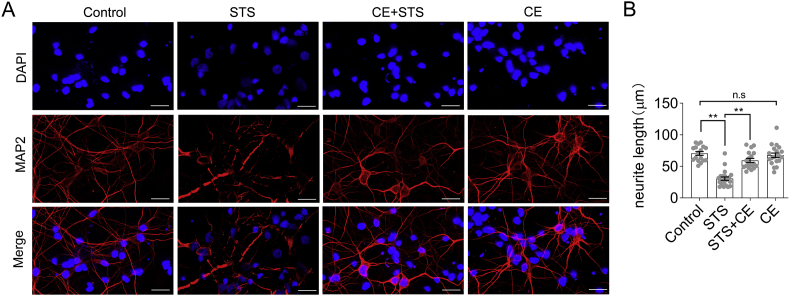
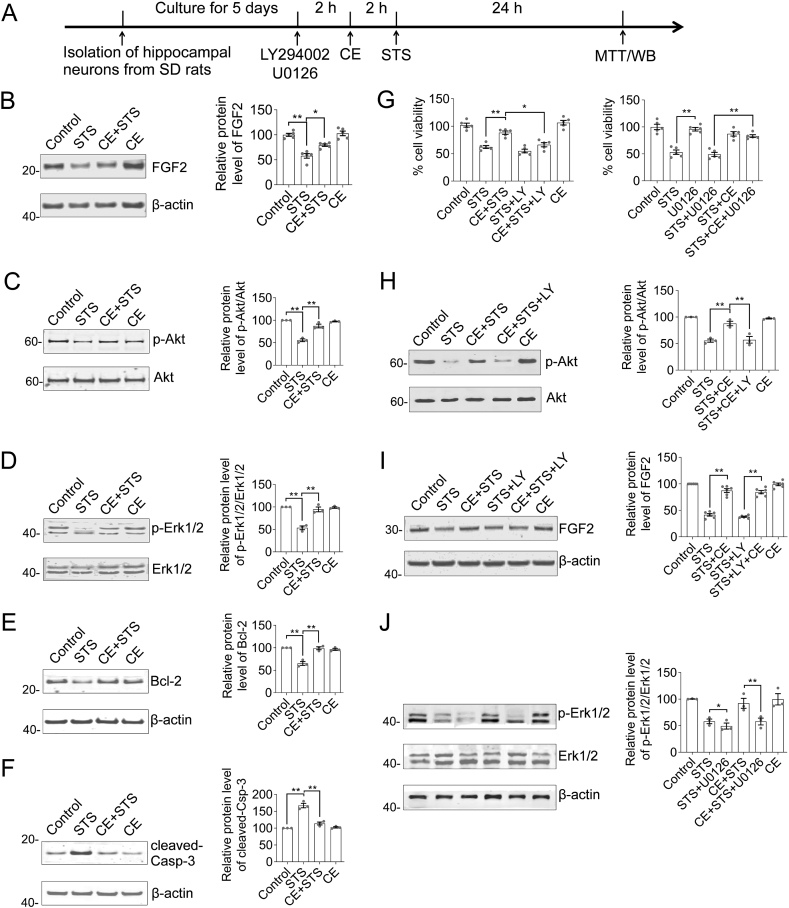
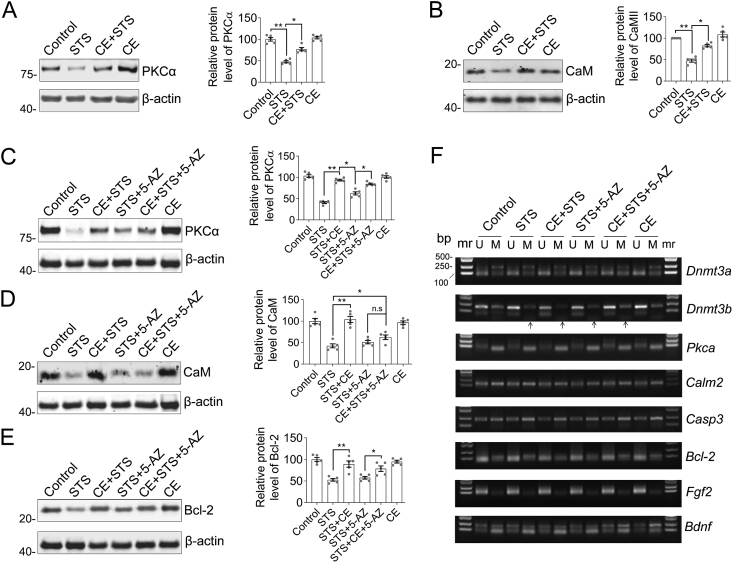
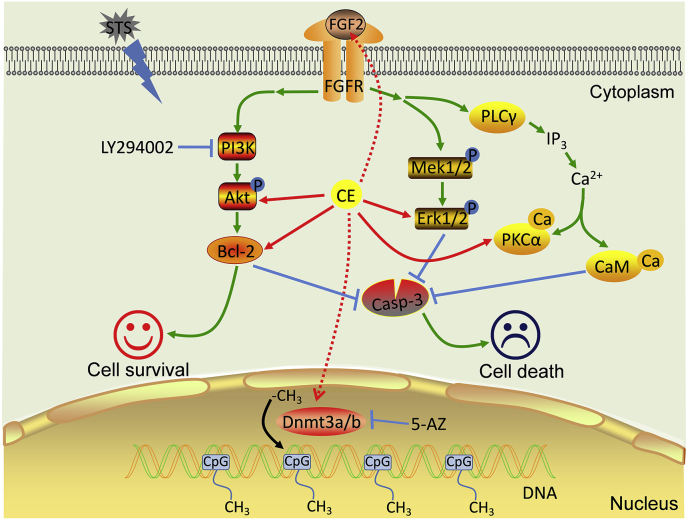
We previously demonstrated the antioxidant activity of Coeloglossum viride var. bracteatum extract (CE) in rat cortical neurons and in mice with chemically induced cognitive impairment. In this work, we established a staurosporine (STS)-induced toxicity model to decipher the neuroprotective mechanisms of CE. We found that CE protected cell viability and neurite integrity in STS-induced toxicity by restoring the levels of FGF2 and its associated PI3K/Akt signaling axis. LY294002, a pan-inhibitor of PI3K, antagonized the activity of CE, although its-mediated restoration of FGF2 was unaffected. In addition, CE restored levels of Bcl-2/Caspase-3, PKCα/CaM pathway, and Dnmt3a and Dnmt3b, two methyltransferases that contribute to de novo DNA methylation. The Dnmts inhibitor 5-azacytidine impaired CE-mediated restoration of Dnmt3 or CaM, as well as the transition of DNA methylation status on the Dnmt3 promoter. These results reveal potential mechanisms that could facilitate the study and application of CE as a neuroprotective agent.
Keywords: CE; Dnmt3a; Dnmt3b; FGF2; PI3K/Akt; Staurosporine.
© 2021 The Author(s).
Conflict of interest statement
The authors declare no conflict of interest.
Figures

References
-
- Ford-Perriss M., Abud H., Murphy M. Fibroblast growth factors in the developing central nervous system. Clin. Exp. Pharmacol. Physiol. 2001;28:493–503. - PubMed
LinkOut - more resources
Full Text Sources
Research Materials
