Parallel adaptation in autopolyploid Arabidopsis arenosa is dominated by repeated recruitment of shared alleles
- PMID: 34404804
- PMCID: PMC8370997
- DOI: 10.1038/s41467-021-25256-5
Parallel adaptation in autopolyploid Arabidopsis arenosa is dominated by repeated recruitment of shared alleles
Abstract
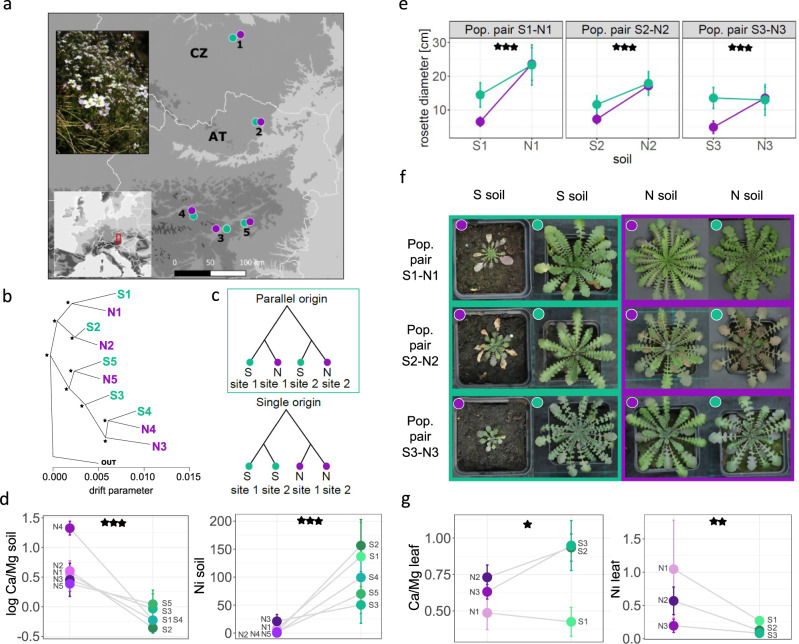
Relative contributions of pre-existing vs de novo genomic variation to adaptation are poorly understood, especially in polyploid organisms. We assess this in high resolution using autotetraploid Arabidopsis arenosa, which repeatedly adapted to toxic serpentine soils that exhibit skewed elemental profiles. Leveraging a fivefold replicated serpentine invasion, we assess selection on SNPs and structural variants (TEs) in 78 resequenced individuals and discover significant parallelism in candidate genes involved in ion homeostasis. We further model parallel selection and infer repeated sweeps on a shared pool of variants in nearly all these loci, supporting theoretical expectations. A single striking exception is represented by TWO PORE CHANNEL 1, which exhibits convergent evolution from independent de novo mutations at an identical, otherwise conserved site at the calcium channel selectivity gate. Taken together, this suggests that polyploid populations can rapidly adapt to environmental extremes, calling on both pre-existing variation and novel polymorphisms.
© 2021. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures