

Neuronal odor coding in the larval sensory cone of Anopheles coluzzii: Complex responses from a simple system
- PMID: 34407405
- PMCID: PMC8420959
- DOI: 10.1016/j.celrep.2021.109555
Neuronal odor coding in the larval sensory cone of Anopheles coluzzii: Complex responses from a simple system
Abstract
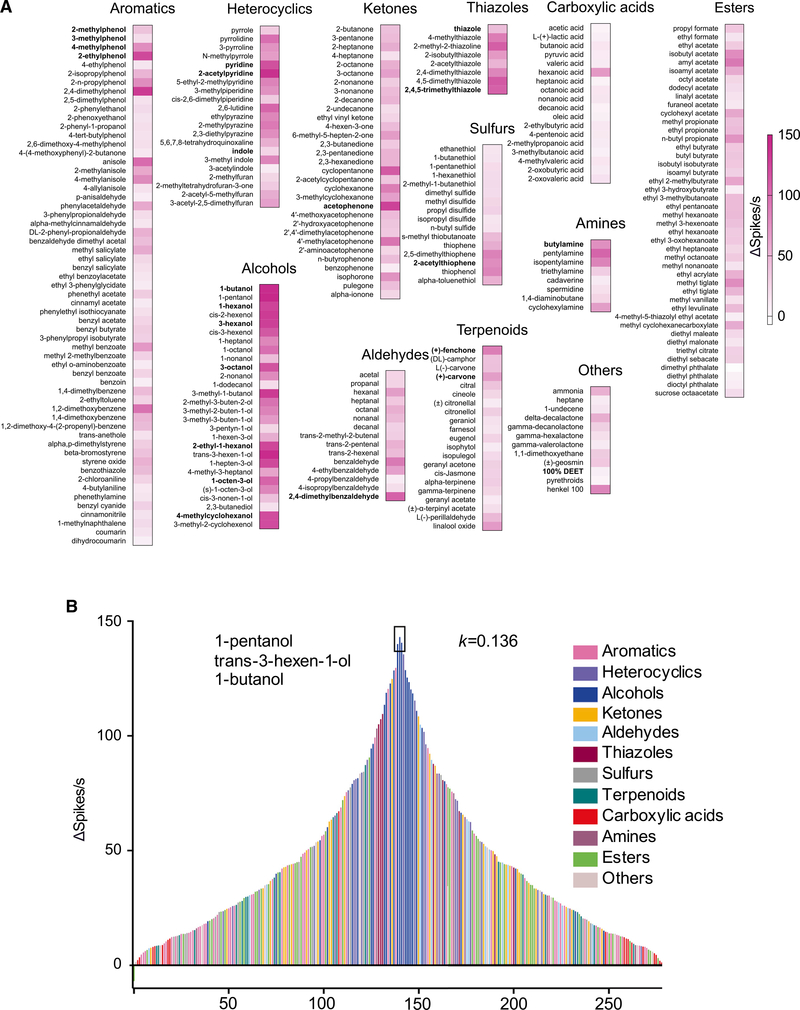
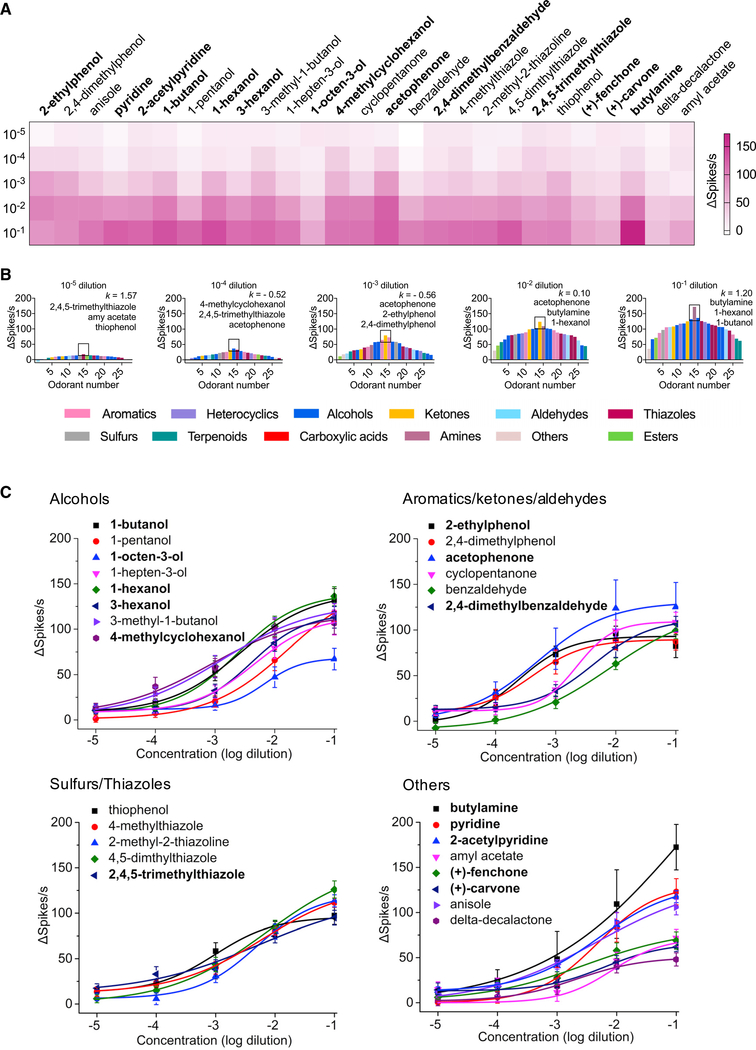
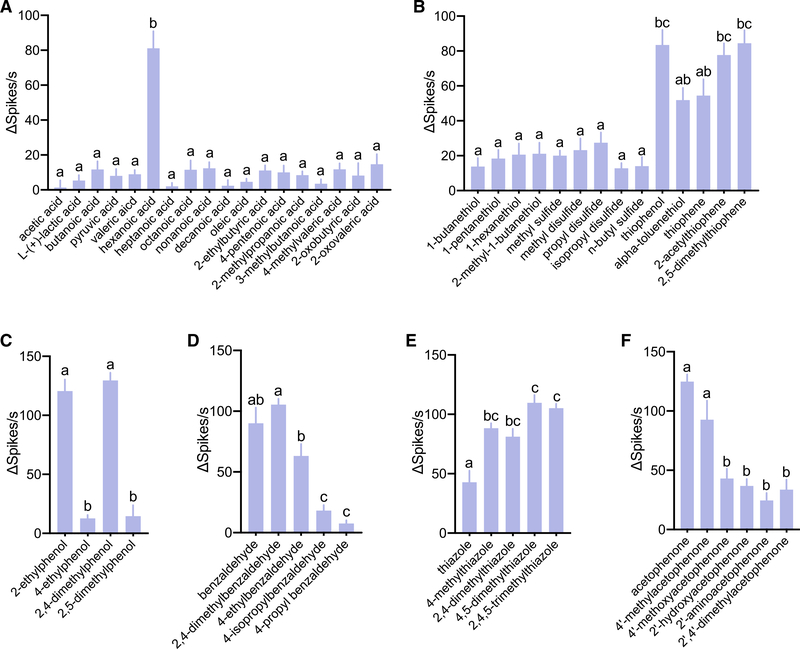
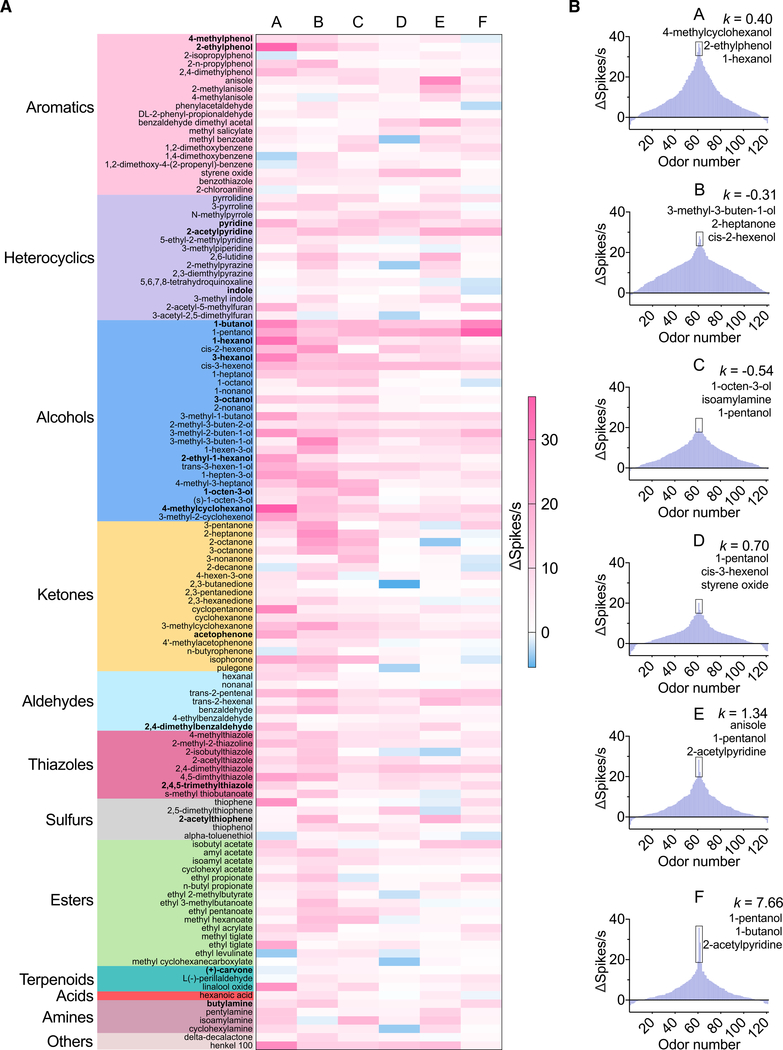
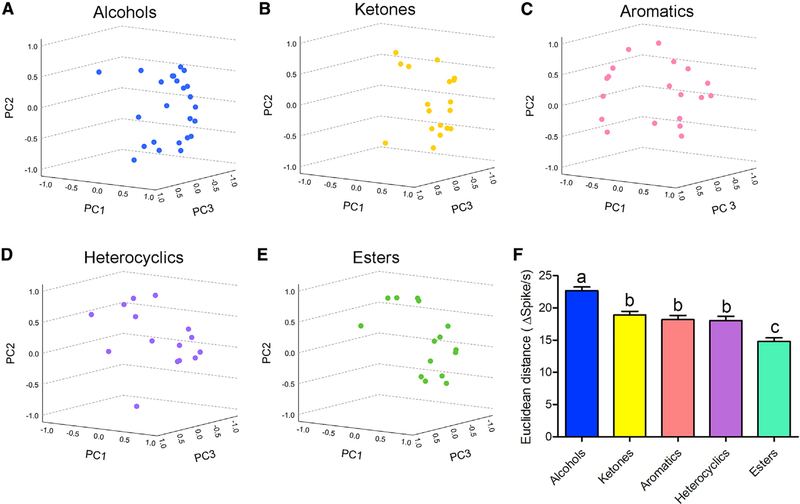
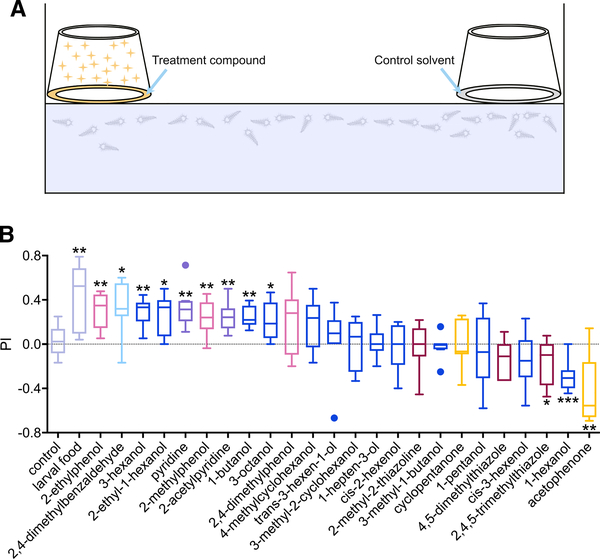
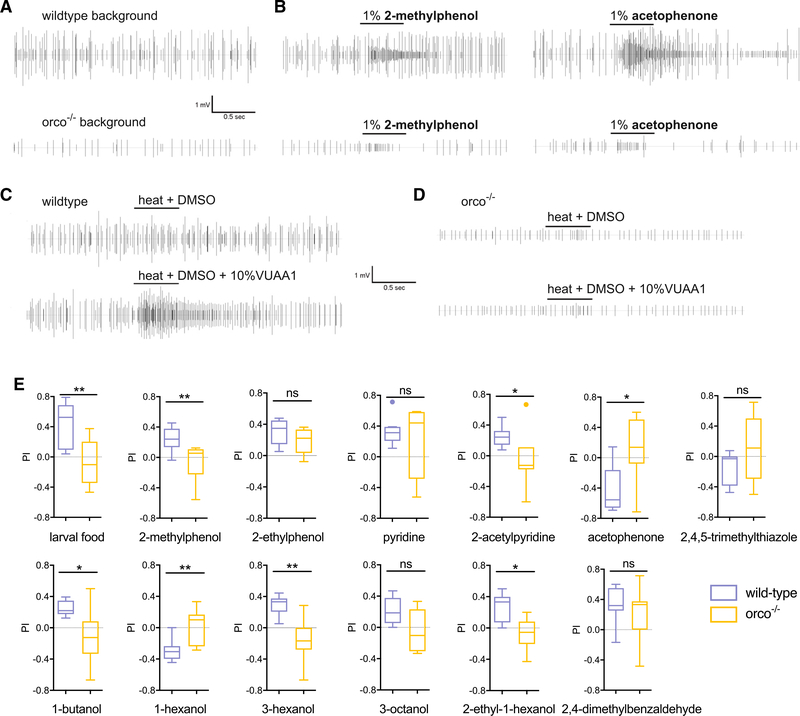
Anopheles mosquitoes are the sole vectors of malaria. Although adult females are directly responsible for disease transmission and accordingly have been extensively studied, the survival of pre-adult larval stages is vital. Mosquito larvae utilize a spectrum of chemosensory and other cues to navigate their aquatic habitats to avoid predators and search for food. Here we examine larval olfactory responses, in which the peripheral components are associated with the antennal sensory cone. Larval behavior and sensory cone responses to volatile stimuli in Anopheles coluzzii demonstrate the sensory cone is particularly tuned to alcohols, thiazoles, and heterocyclics, and these responses can be assigned to discrete groups of sensory cone neurons with distinctive profiles. These studies reveal that the anopheline larvae actively sample volatile odors above their aquatic habitats via a highly sophisticated olfactory system that is sensitive to a broad range of compounds with significant behavioral relevance.
Keywords: Anopheles coluzzii; antennal sensory cone; behavior; electrophysiology; larval olfactory system.
Copyright © 2021 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
References
-
- Audrain B, Farag MA, Ryu CM, and Ghigo JM (2015). Role of bacterial volatile compounds in bacterial biology. FEMS Microbiol. Rev. 39, 222–233. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
