

Coordination of tumor growth and host wasting by tumor-derived Upd3
- PMID: 34407411
- PMCID: PMC8410949
- DOI: 10.1016/j.celrep.2021.109553
Coordination of tumor growth and host wasting by tumor-derived Upd3
Abstract
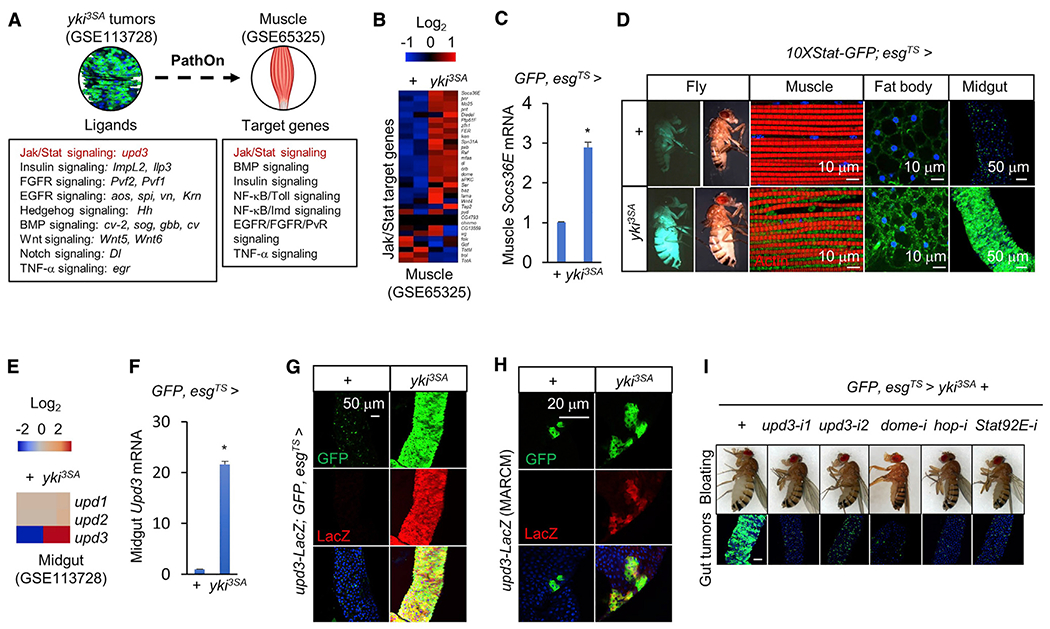
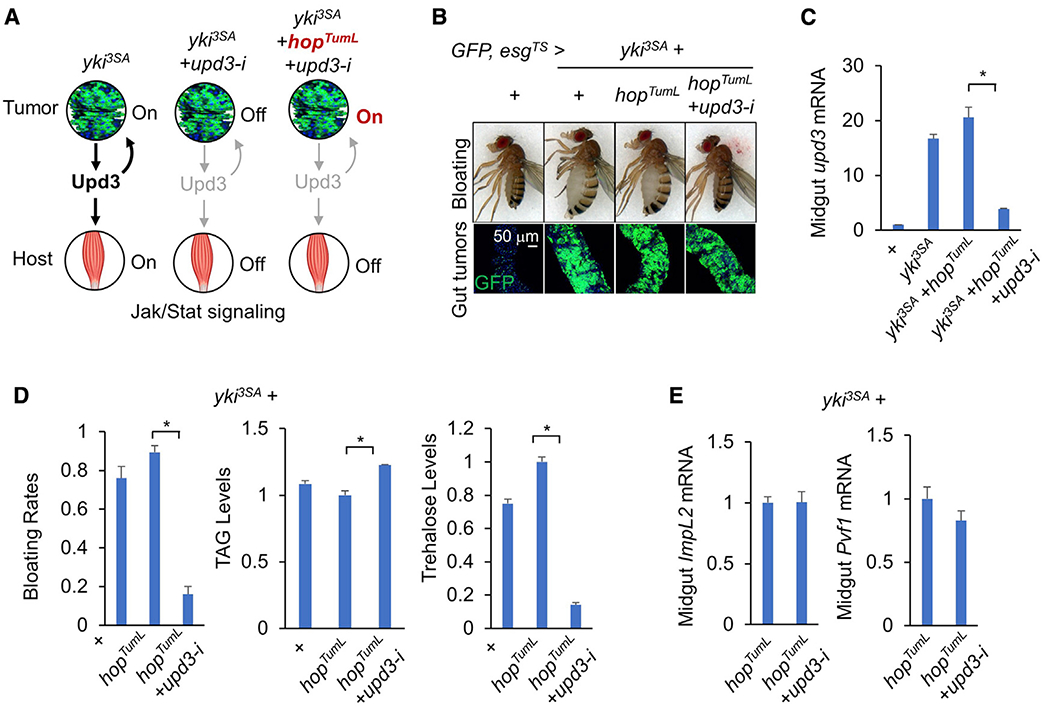
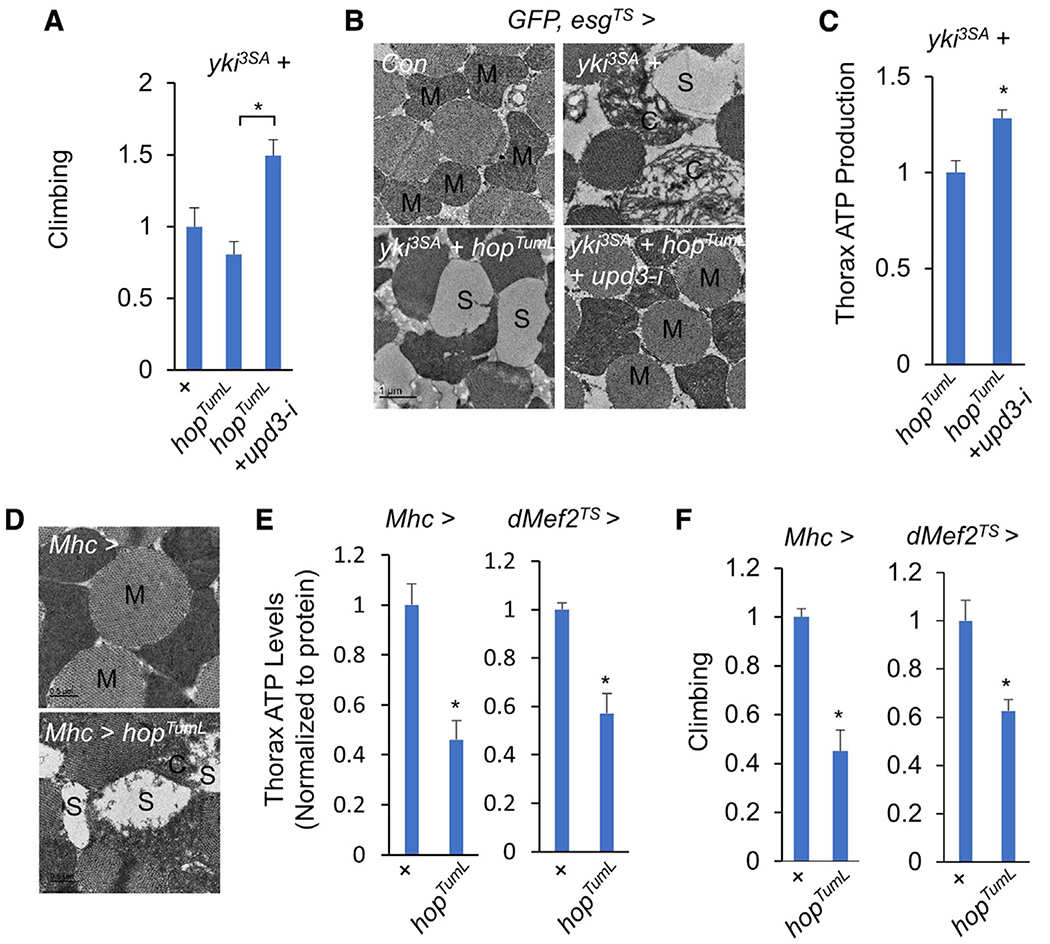
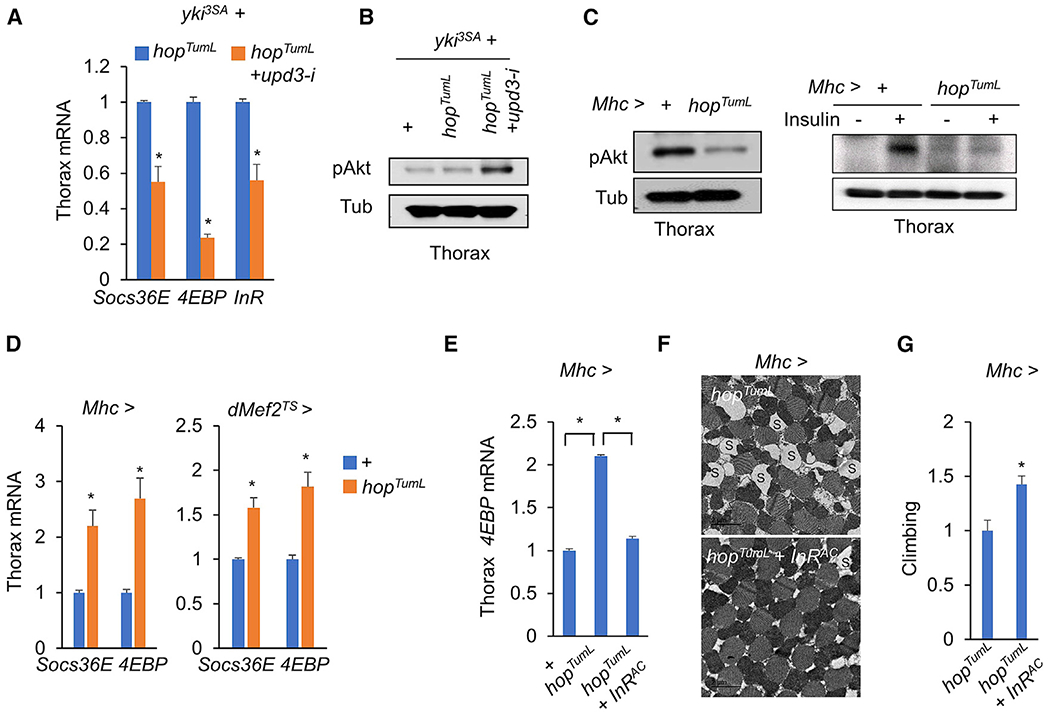
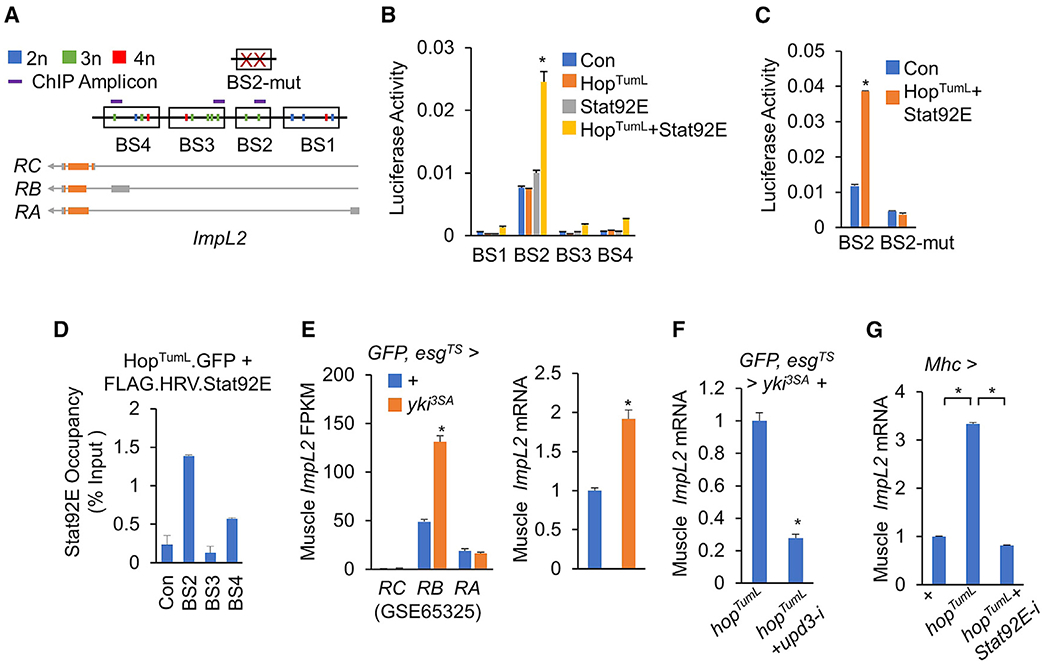
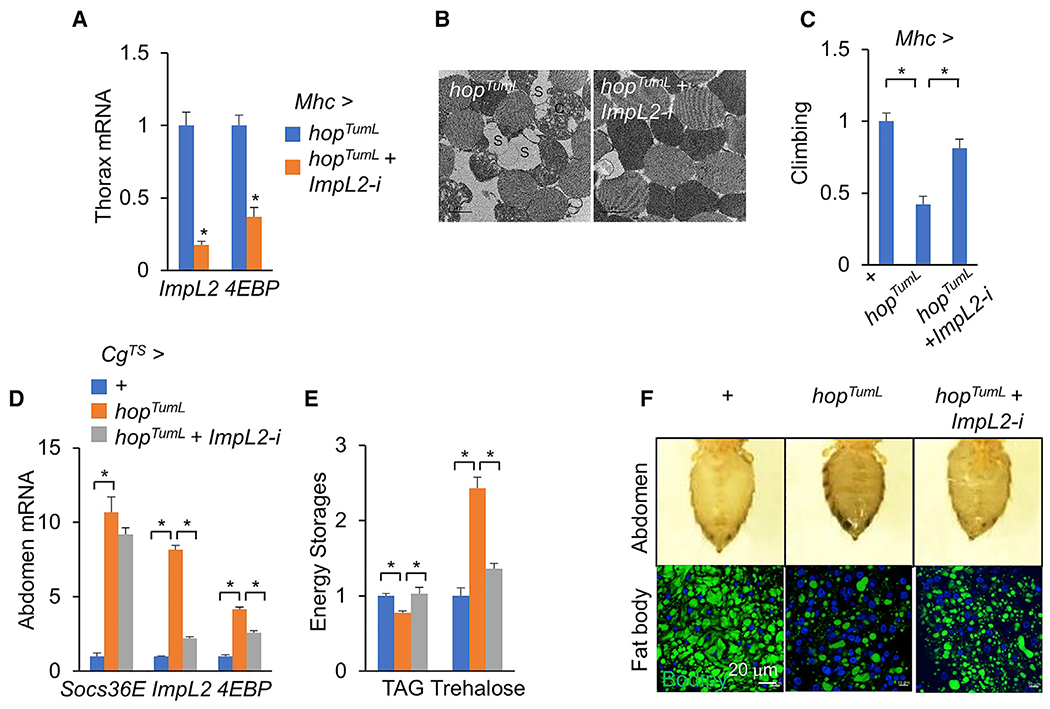
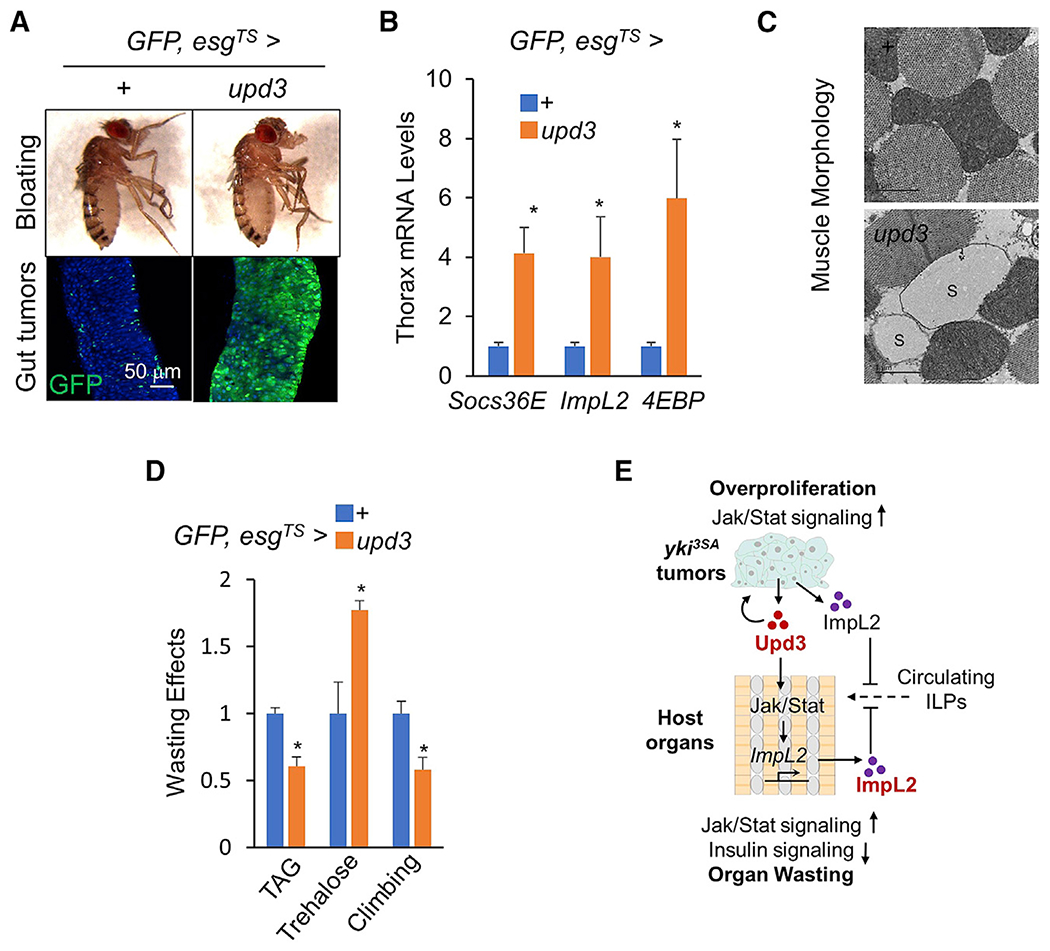
yki-induced gut tumors in Drosophila are associated with host wasting, including muscle dysfunction, lipid loss, and hyperglycemia, a condition reminiscent of human cancer cachexia. We previously used this model to identify tumor-derived ligands that contribute to host wasting. To identify additional molecular networks involved in host-tumor interactions, we develop PathON, a web-based tool analyzing the major signaling pathways in Drosophila, and uncover the Upd3/Jak/Stat axis as an important modulator. We find that yki-gut tumors secrete Upd3 to promote self-overproliferation and enhance Jak/Stat signaling in host organs to cause wasting, including muscle dysfunction, lipid loss, and hyperglycemia. We further reveal that Upd3/Jak/Stat signaling in the host organs directly triggers the expression of ImpL2, an antagonistic binding protein for insulin-like peptides, to impair insulin signaling and energy balance. Altogether, our results demonstrate that yki-gut tumors produce a Jak/Stat pathway ligand, Upd3, that regulates both self-growth and host wasting.
Keywords: Drosophila; ImpL2; Jak/Stat signaling; Upd3; cancer cachexia; lipid loss; muscle dysfunction; tumor-induced wasting; yki.
Copyright © 2021 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
Similar articles
-
The UPD3 cytokine couples environmental challenge and intestinal stem cell division through modulation of JAK/STAT signaling in the stem cell microenvironment.Dev Biol. 2013 Jan 15;373(2):383-93. doi: 10.1016/j.ydbio.2012.10.023. Epub 2012 Oct 27. Dev Biol. 2013. PMID: 23110761 Free PMC article.
-
Macrophage-like Blood Cells Are Involved in Inter-Tissue Communication to Activate JAK/STAT Signaling, Inducing Antitumor Turandot Proteins in Drosophila Fat Body via the TNF-JNK Pathway.Int J Mol Sci. 2024 Dec 6;25(23):13110. doi: 10.3390/ijms252313110. Int J Mol Sci. 2024. PMID: 39684820 Free PMC article.
-
Subpopulation of Macrophage-Like Plasmatocytes Attenuates Systemic Growth via JAK/STAT in the Drosophila Fat Body.Front Immunol. 2020 Jan 31;11:63. doi: 10.3389/fimmu.2020.00063. eCollection 2020. Front Immunol. 2020. PMID: 32082322 Free PMC article.
-
Novel studies on Drosophila melanogaster model reveal the roles of JNK-Jak/STAT axis and intestinal microbiota in insulin resistance.J Drug Target. 2023 Mar;31(3):261-268. doi: 10.1080/1061186X.2022.2144869. Epub 2022 Nov 10. J Drug Target. 2023. PMID: 36343203 Review.
-
Drosophila Jak/STAT Signaling: Regulation and Relevance in Human Cancer and Metastasis.Int J Mol Sci. 2018 Dec 14;19(12):4056. doi: 10.3390/ijms19124056. Int J Mol Sci. 2018. PMID: 30558204 Free PMC article. Review.
Cited by
-
A tumor-secreted protein utilizes glucagon release to cause host wasting.Cell Discov. 2025 Feb 10;11(1):11. doi: 10.1038/s41421-024-00762-0. Cell Discov. 2025. PMID: 39924534 Free PMC article.
-
Body composition and lung cancer-associated cachexia in TRACERx.Nat Med. 2023 Apr;29(4):846-858. doi: 10.1038/s41591-023-02232-8. Epub 2023 Apr 12. Nat Med. 2023. PMID: 37045997 Free PMC article.
-
Cachexia: A systemic consequence of progressive, unresolved disease.Cell. 2023 Apr 27;186(9):1824-1845. doi: 10.1016/j.cell.2023.03.028. Cell. 2023. PMID: 37116469 Free PMC article. Review.
-
A comprehensive review of animal models for cancer cachexia: Implications for translational research.Genes Dis. 2023 Sep 13;11(6):101080. doi: 10.1016/j.gendis.2023.101080. eCollection 2024 Nov. Genes Dis. 2023. PMID: 39220755 Free PMC article. Review.
-
FlyRNAi.org 2025 update-expanded resources for new technologies and species.Nucleic Acids Res. 2025 Jan 6;53(D1):D958-D965. doi: 10.1093/nar/gkae917. Nucleic Acids Res. 2025. PMID: 39435987 Free PMC article.
References
-
- Acharyya S, and Guttridge DC, (2007). Cancer cachexia signaling pathways continue to emerge yet much still points to the proteasome. Clin. Cancer Res 13, 1356–1361. - PubMed
-
- Bach EA, Ekas LA, Ayala-Camargo A, Flaherty MS, Lee H, Perrimon N, and Baeg GH, (2007). GFP reporters detect the activation of the Drosophila JAK/STAT pathway in vivo. Gene Expr. Patterns 7, 323–331. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
