

Reprogrammed lipid metabolism protects inner nuclear membrane against unsaturated fat
- PMID: 34407429
- PMCID: PMC8480995
- DOI: 10.1016/j.devcel.2021.07.018
Reprogrammed lipid metabolism protects inner nuclear membrane against unsaturated fat
Abstract
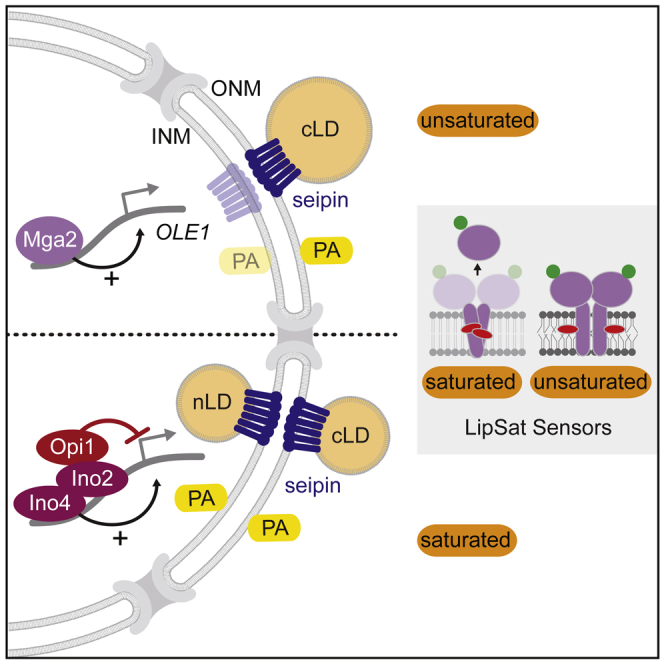
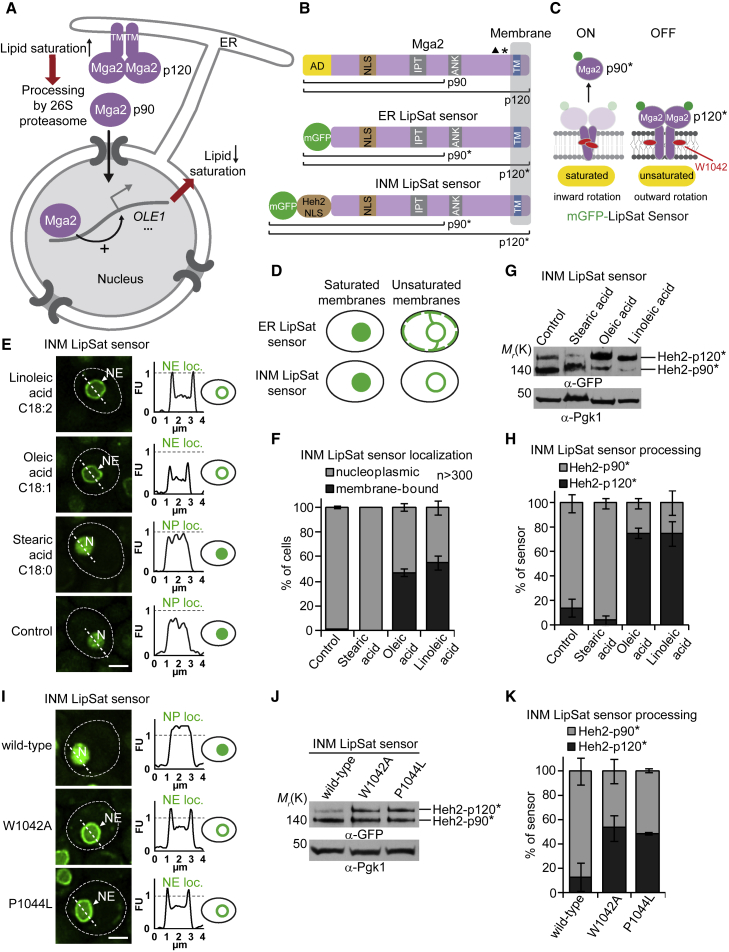
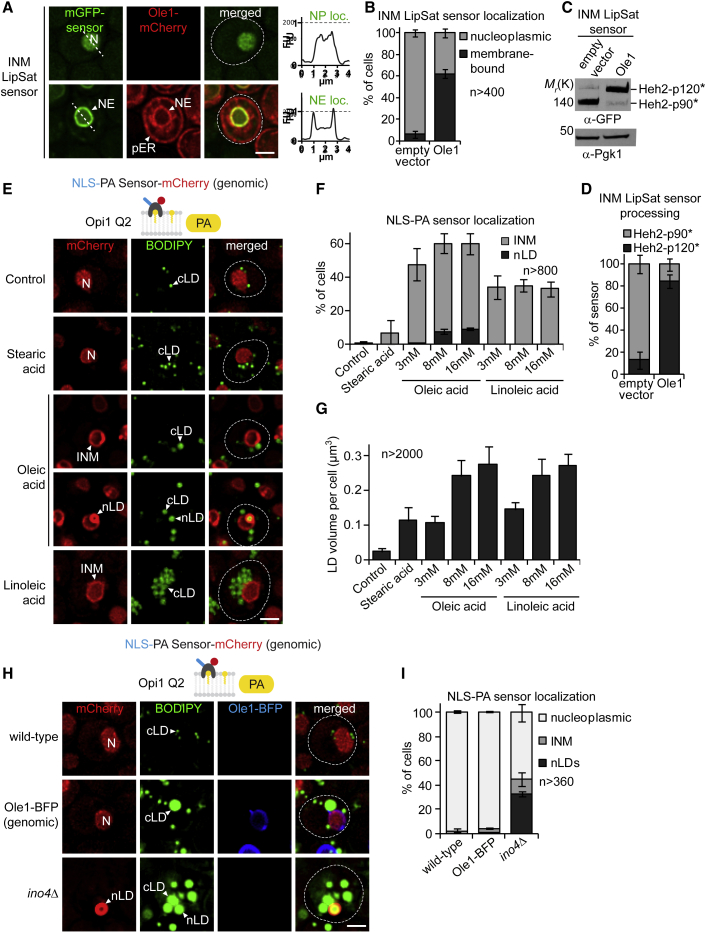
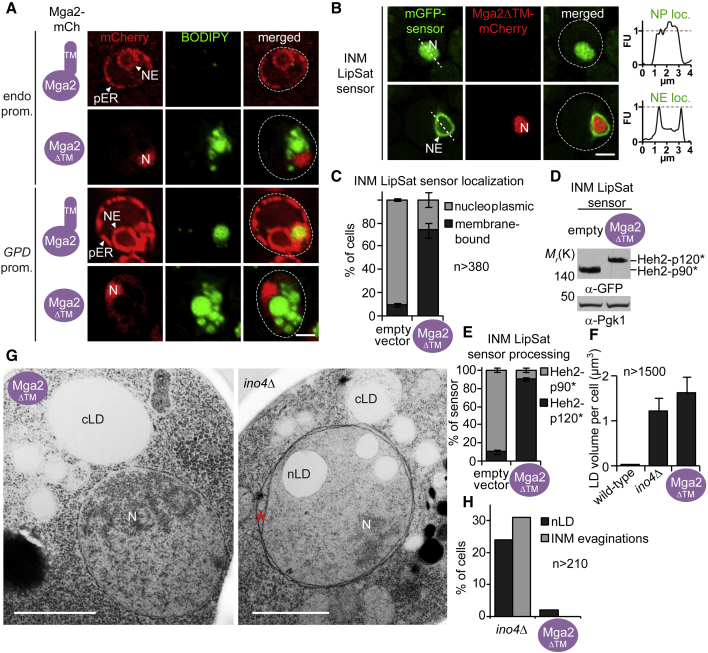
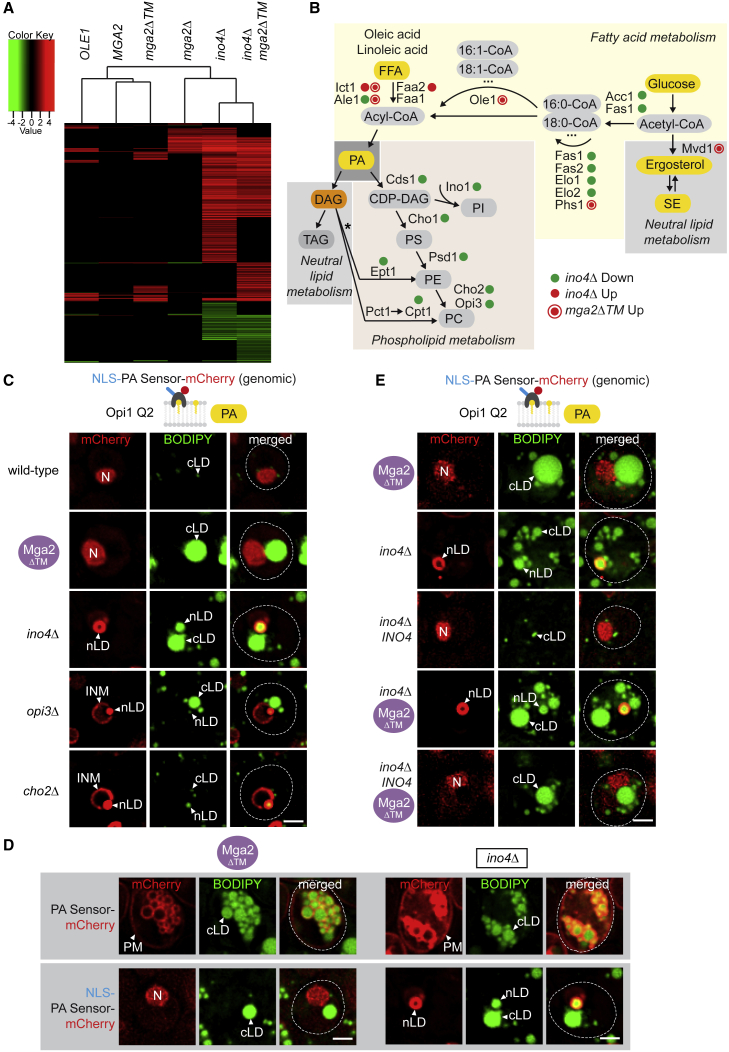
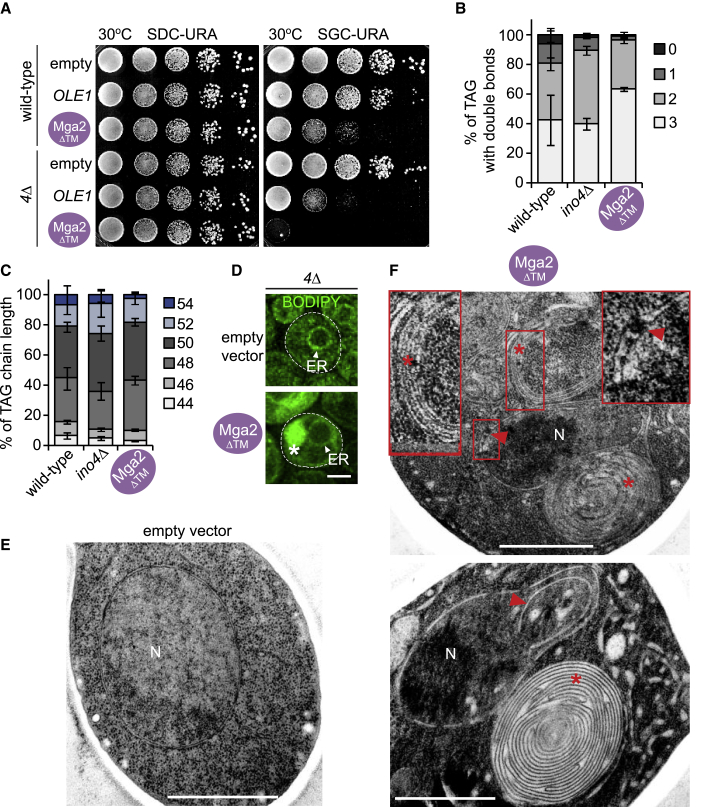
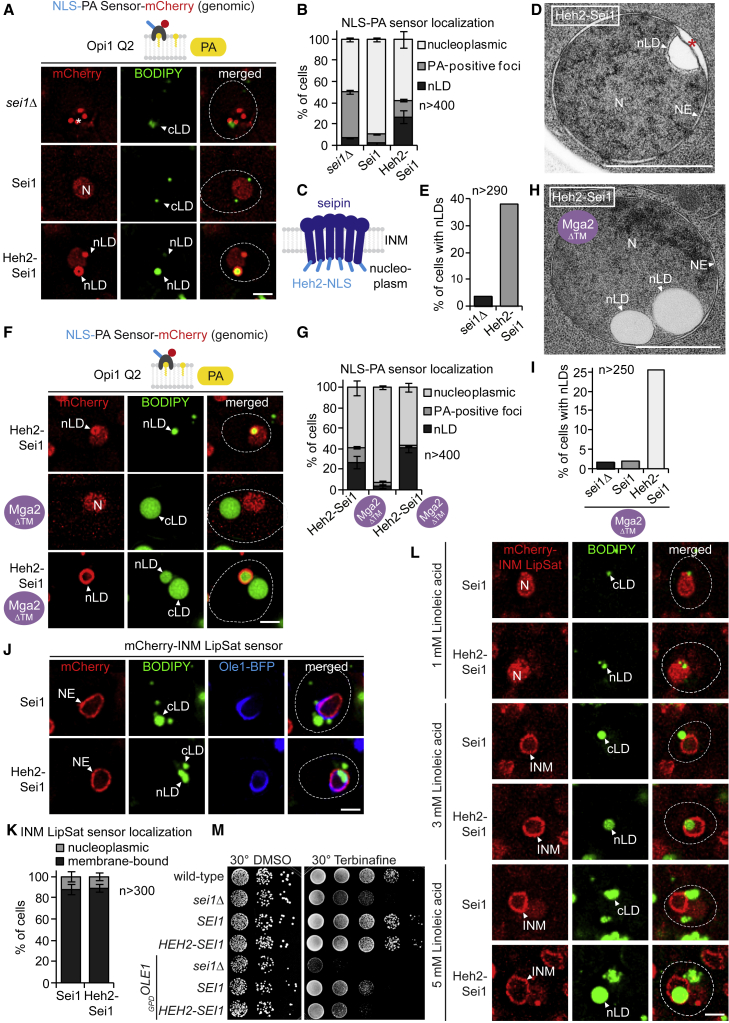
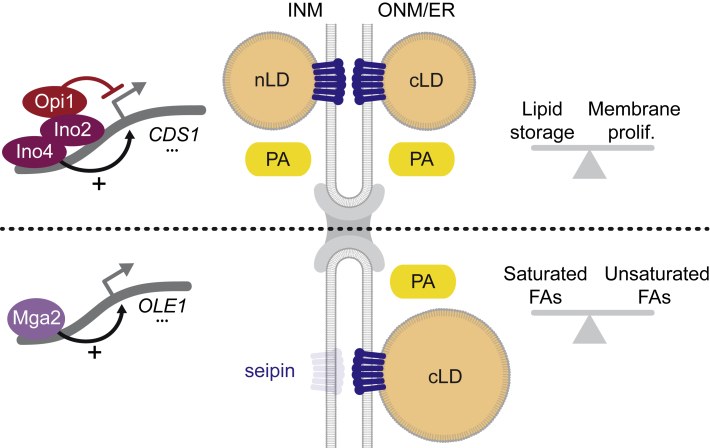
The cell nucleus is surrounded by a double membrane. The lipid packing and viscosity of membranes is critical for their function and is tightly controlled by lipid saturation. Circuits regulating the lipid saturation of the outer nuclear membrane (ONM) and contiguous endoplasmic reticulum (ER) are known. However, how lipid saturation is controlled in the inner nuclear membrane (INM) has remained enigmatic. Using INM biosensors and targeted genetic manipulations, we show that increased lipid unsaturation causes a reprogramming of lipid storage metabolism across the nuclear envelope (NE). Cells induce lipid droplet (LD) formation specifically from the distant ONM/ER, whereas LD formation at the INM is suppressed. In doing so, unsaturated fatty acids are shifted away from the INM. We identify the transcription circuits that topologically reprogram LD synthesis and identify seipin and phosphatidic acid as critical effectors. Our study suggests a detoxification mechanism protecting the INM from excess lipid unsaturation.
Keywords: Mga2/Ole1; endoplasmic reticulum; inner nuclear membrane; lipid biosensors; lipid droplets; lipid metabolism; nuclear envelope; phosphatidic acid; seipin; unsaturated fatty acids.
Copyright © 2021 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
Comment in
-
Nuclear detox of unsaturated fat.Dev Cell. 2021 Sep 27;56(18):2539-2541. doi: 10.1016/j.devcel.2021.09.002. Dev Cell. 2021. PMID: 34582769
References
-
- Bigay J., Antonny B. Curvature, lipid packing, and electrostatics of membrane organelles: defining cellular territories in determining specificity. Dev. Cell. 2012;23:886–895. - PubMed
-
- Black P.N., DiRusso C.C. Yeast acyl-CoA synthetases at the crossroads of fatty acid metabolism and regulation. Biochimica et Biophysica Acta (BBA) - Molecular. and. Cell. Biology. of. Lipids. 2007;1771:286–298. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
