

Apolipoprotein M and Sphingosine-1-Phosphate Receptor 1 Promote the Transendothelial Transport of High-Density Lipoprotein
- PMID: 34407633
- PMCID: PMC8458249
- DOI: 10.1161/ATVBAHA.121.316725
Apolipoprotein M and Sphingosine-1-Phosphate Receptor 1 Promote the Transendothelial Transport of High-Density Lipoprotein
Abstract
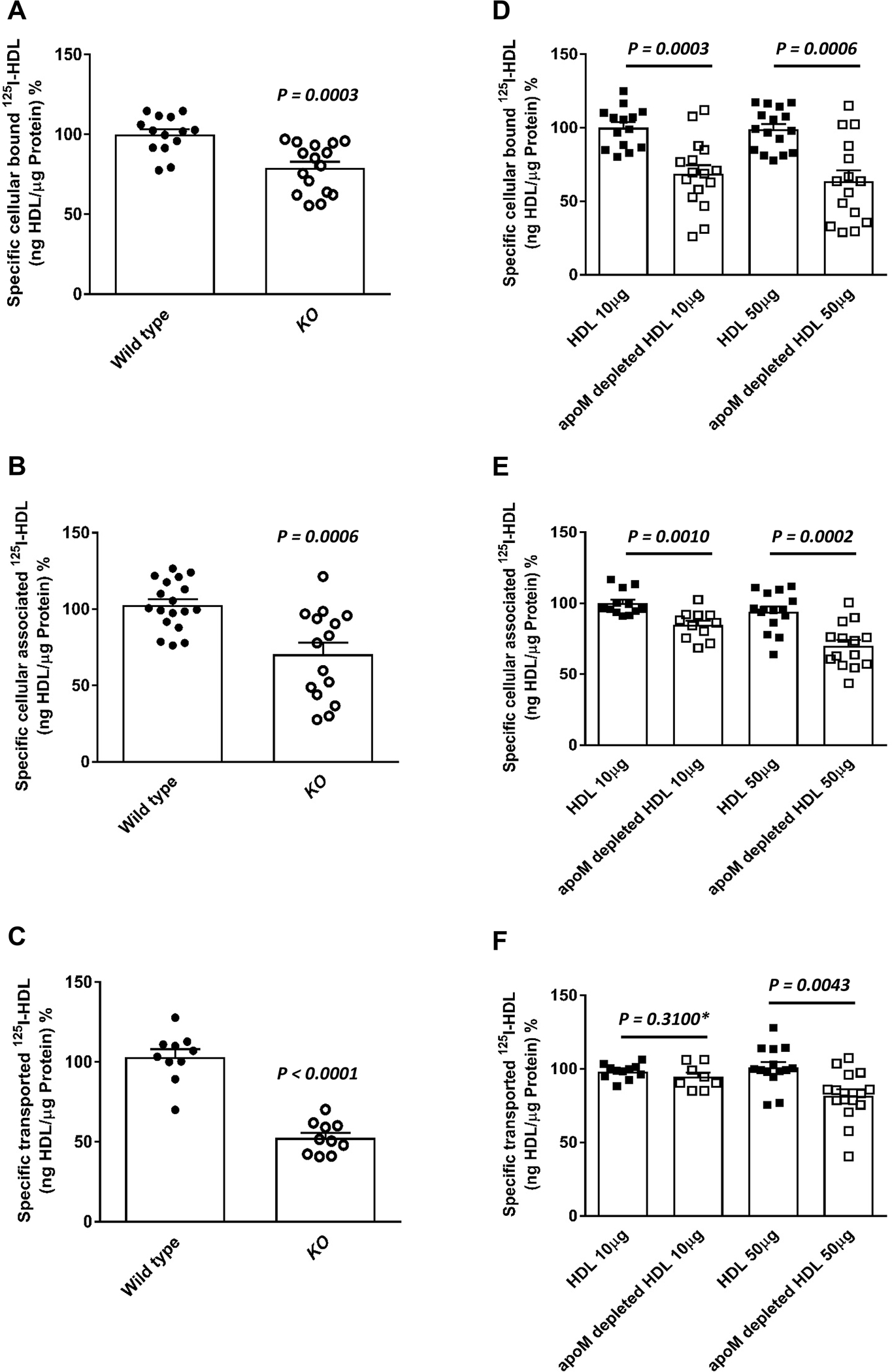
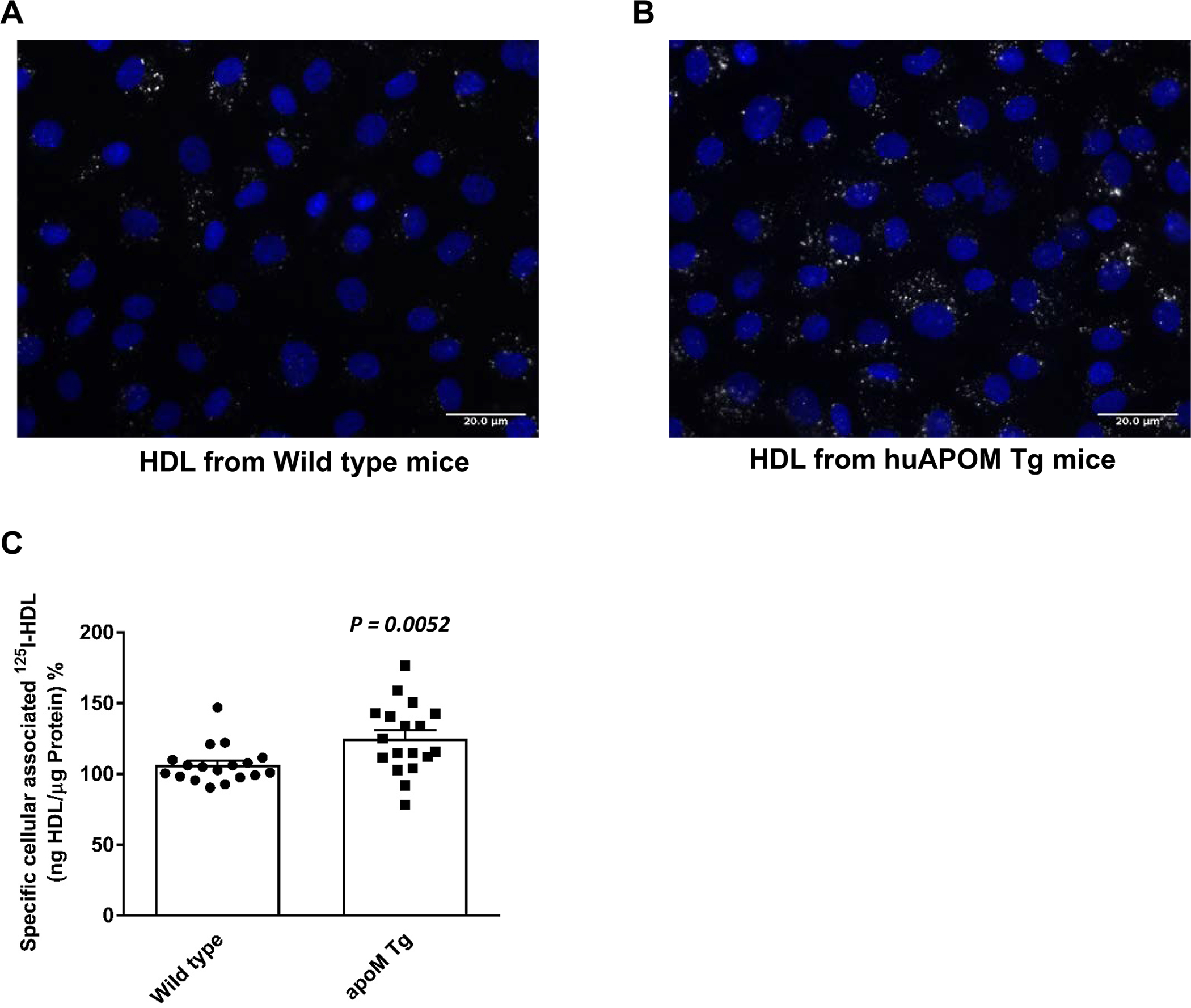
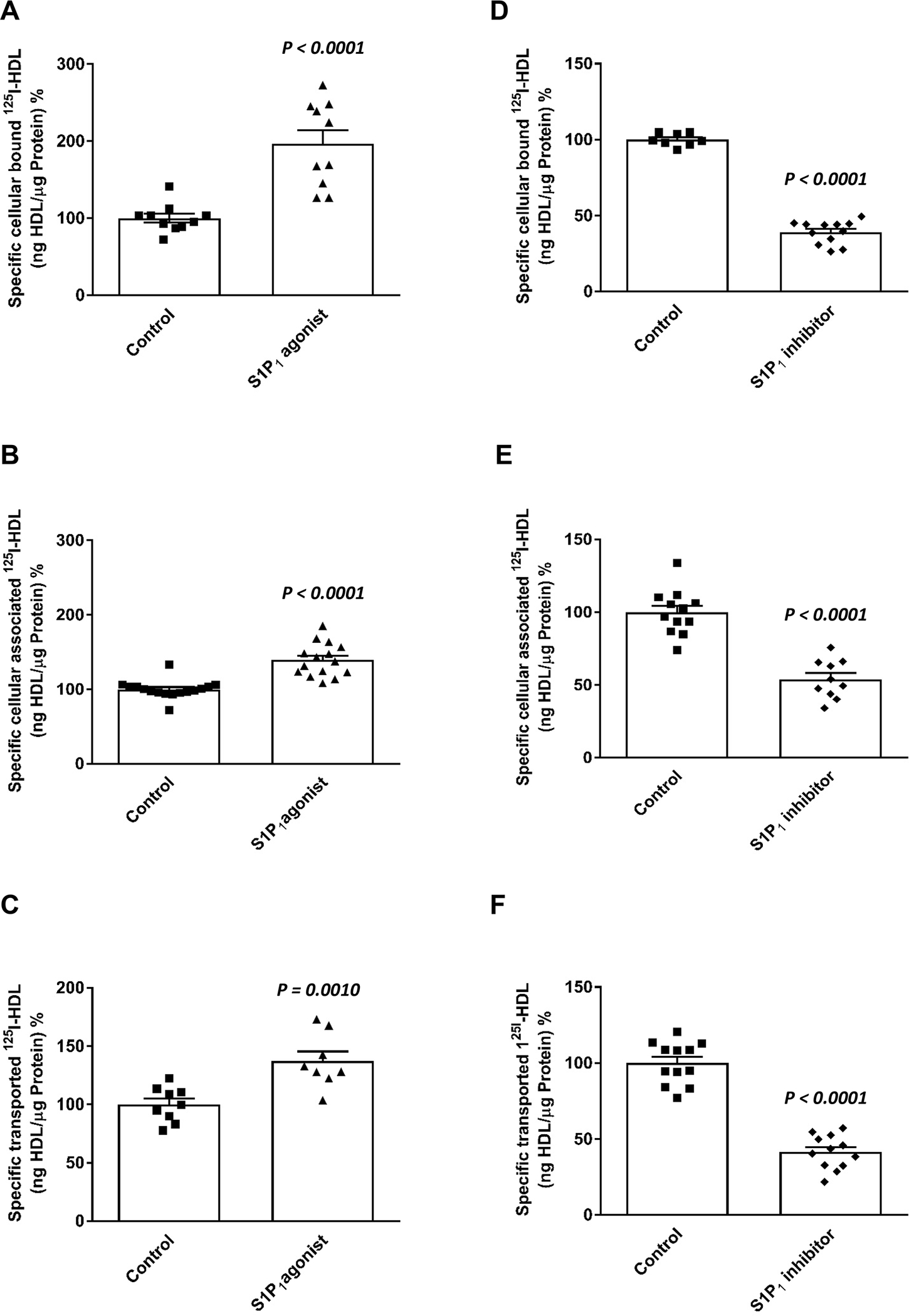
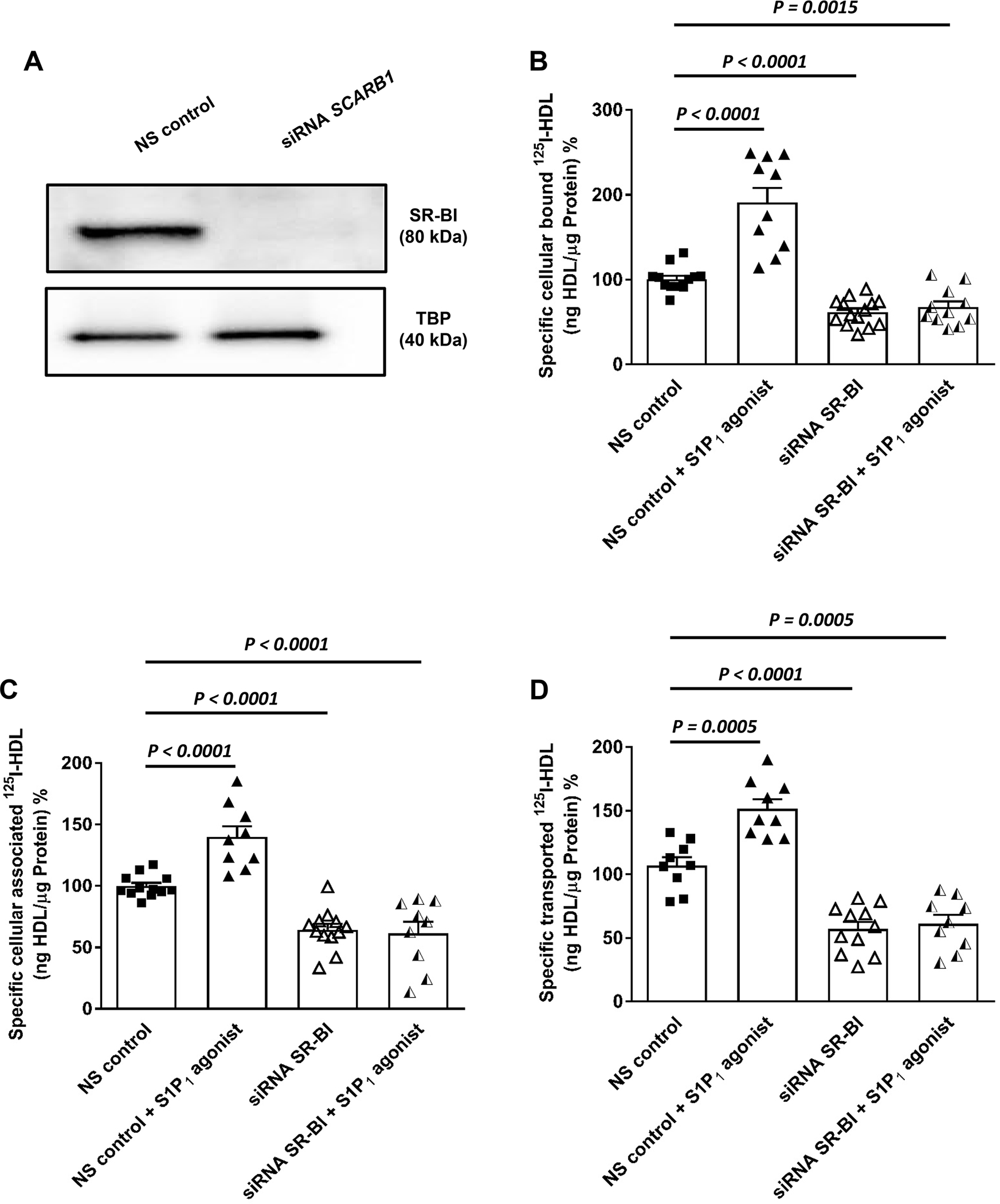
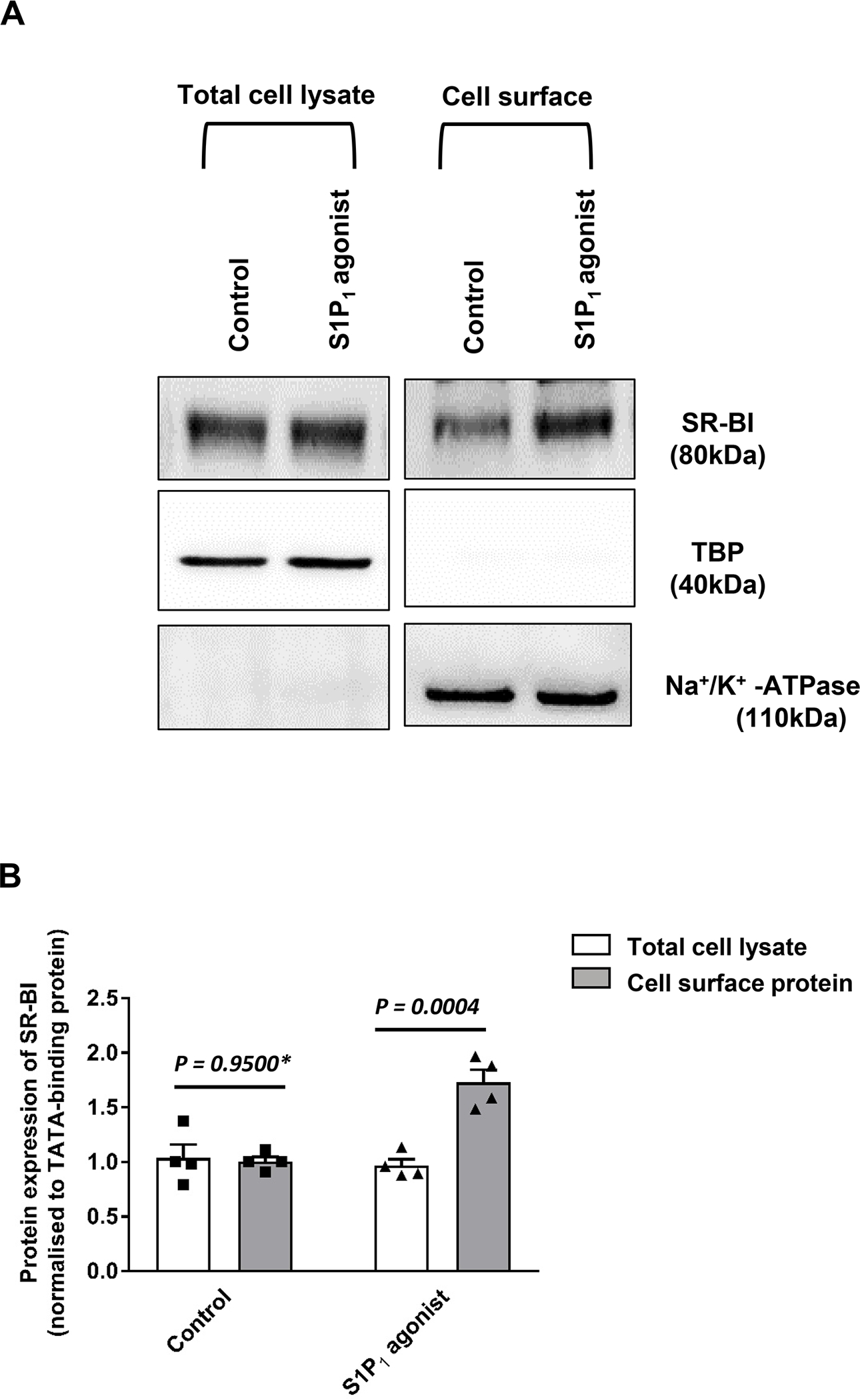
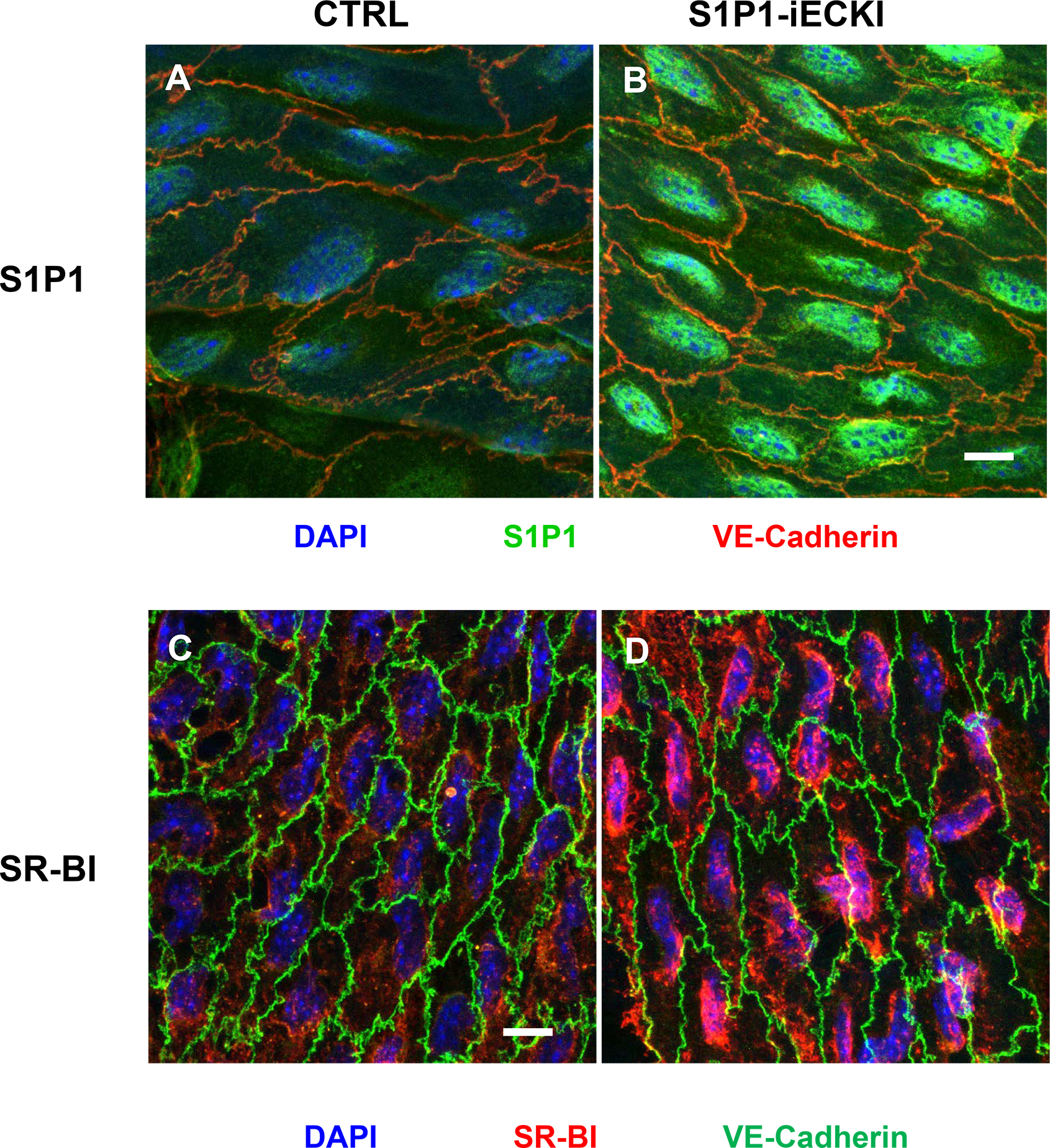
Objective: ApoM enriches S1P (sphingosine-1-phosphate) within HDL (high-density lipoproteins) and facilitates the activation of the S1P1 (S1P receptor type 1) by S1P, thereby preserving endothelial barrier function. Many protective functions exerted by HDL in extravascular tissues raise the question of how S1P regulates transendothelial HDL transport. Approach and Results: HDL were isolated from plasma of wild-type mice, Apom knockout mice, human apoM transgenic mice or humans and radioiodinated to trace its binding, association, and transport by bovine or human aortic endothelial cells. We also compared the transport of fluorescently-labeled HDL or Evans Blue, which labels albumin, from the tail vein into the peritoneal cavity of apoE-haploinsufficient mice with (apoE-haploinsufficient mice with endothelium-specific knockin of S1P1) or without (control mice, ie, apoE-haploinsufficient mice without endothelium-specific knockin of S1P1) endothelium-specific knockin of S1P1. The binding, association, and transport of HDL from Apom knockout mice and human apoM-depleted HDL by bovine aortic endothelial cells was significantly lower than that of HDL from wild-type mice and human apoM-containing HDL, respectively. The binding, uptake, and transport of 125I-HDL by human aortic endothelial cells was increased by an S1P1 agonist but decreased by an S1P1 inhibitor. Silencing of SR-BI (scavenger receptor BI) abrogated the stimulation of 125I-HDL transport by the S1P1 agonist. Compared with control mice, that is, apoE-haploinsufficient mice without endothelium-specific knockin of S1P1, apoE-haploinsufficient mice with endothelium-specific knockin of S1P1 showed decreased transport of Evans Blue but increased transport of HDL from blood into the peritoneal cavity and SR-BI expression in the aortal endothelium. Conclusions: ApoM and S1P1 promote transendothelial HDL transport. Their opposite effect on transendothelial transport of albumin and HDL indicates that HDL passes endothelial barriers by specific mechanisms rather than passive filtration.
Keywords: apolipoprotein; endothelium; lipoprotein; mice; sphingosine-1-phosphate.
Conflict of interest statement
Disclosure
MS is a scientific advisory board member of Alnylam Pharmaceuticals. The other authors have no conflict of interest to disclose with respect to the work described except the funding mentioned above
Figures
References
-
- Annema W, von Eckardstein A. High-density lipoproteins. Multifunctional but vulnerable protections from atherosclerosis. Circ J. 2013;77:2432–2448 - PubMed
-
- Cardner M, Yalcinkaya M, Goetze S, Luca E, Balaz M, Hunjadi M, Hartung J, Shemet A, Krankel N, Radosavljevic S, Keel M, Othman A, Karsai G, Hornemann T, Claassen M, Liebisch G, Carreira E, Ritsch A, Landmesser U, Krutzfeldt J, Wolfrum C, Wollscheid B, Beerenwinkel N, Rohrer L, von Eckardstein A. Structure-function relationships of hdl in diabetes and coronary heart disease. JCI Insight. 2020;5 - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
