

Characterization of the ABC methionine transporter from Neisseria meningitidis reveals that lipidated MetQ is required for interaction
- PMID: 34409939
- PMCID: PMC8416018
- DOI: 10.7554/eLife.69742
Characterization of the ABC methionine transporter from Neisseria meningitidis reveals that lipidated MetQ is required for interaction
Abstract
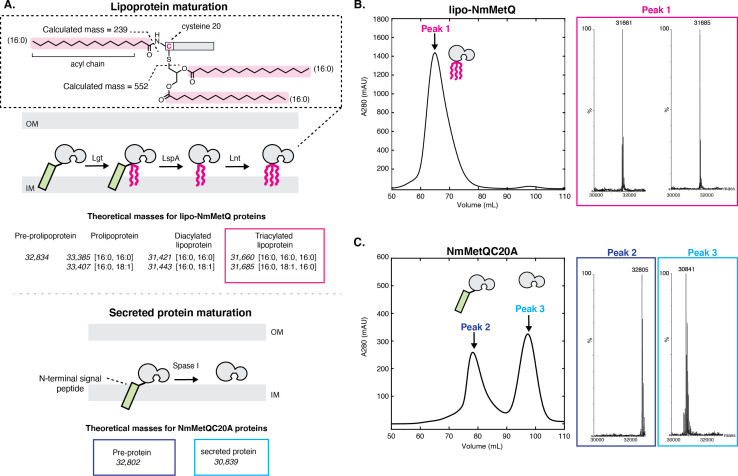
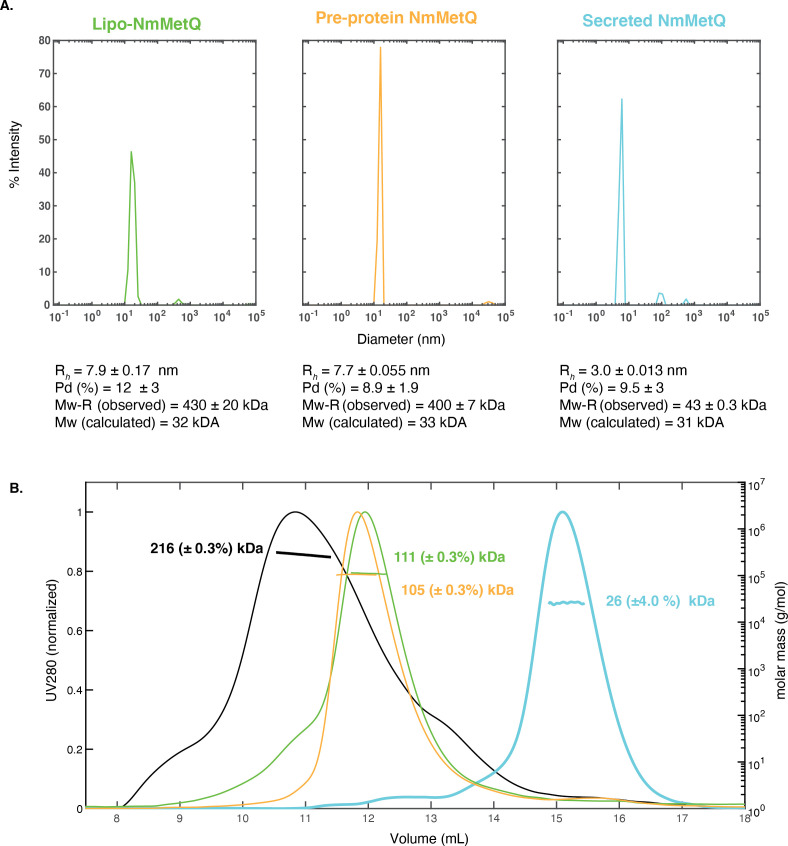
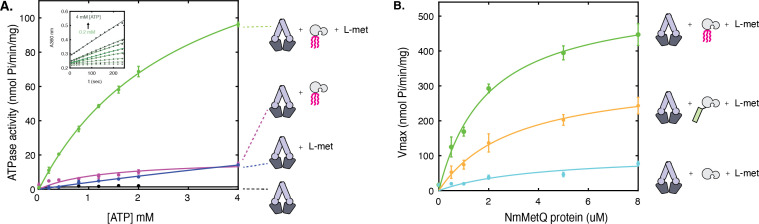
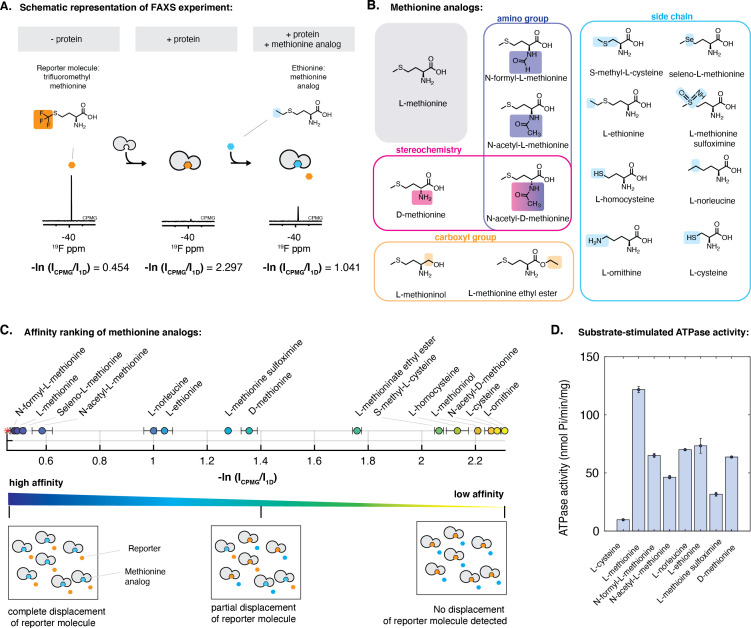
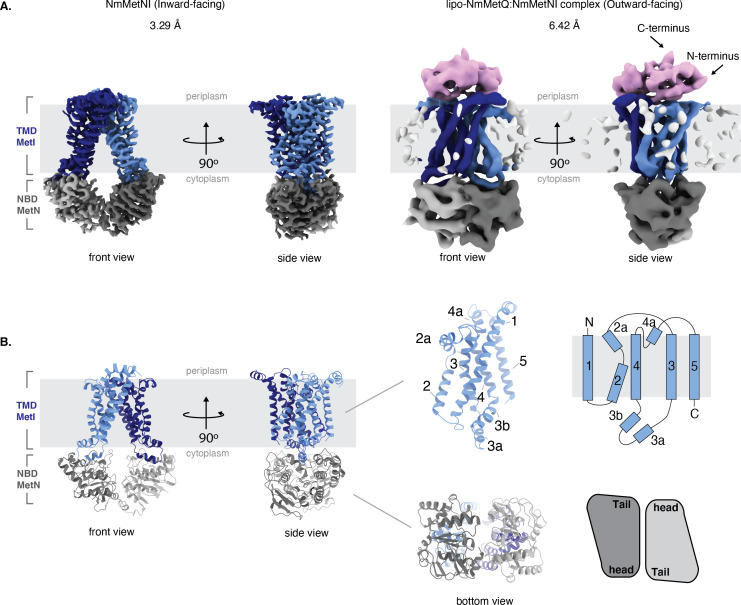
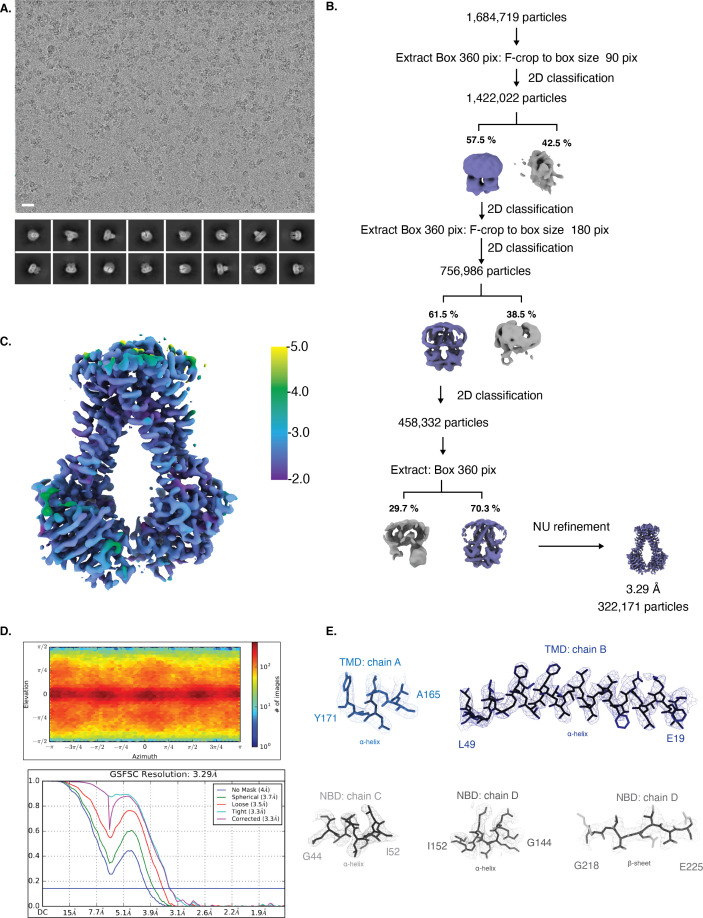
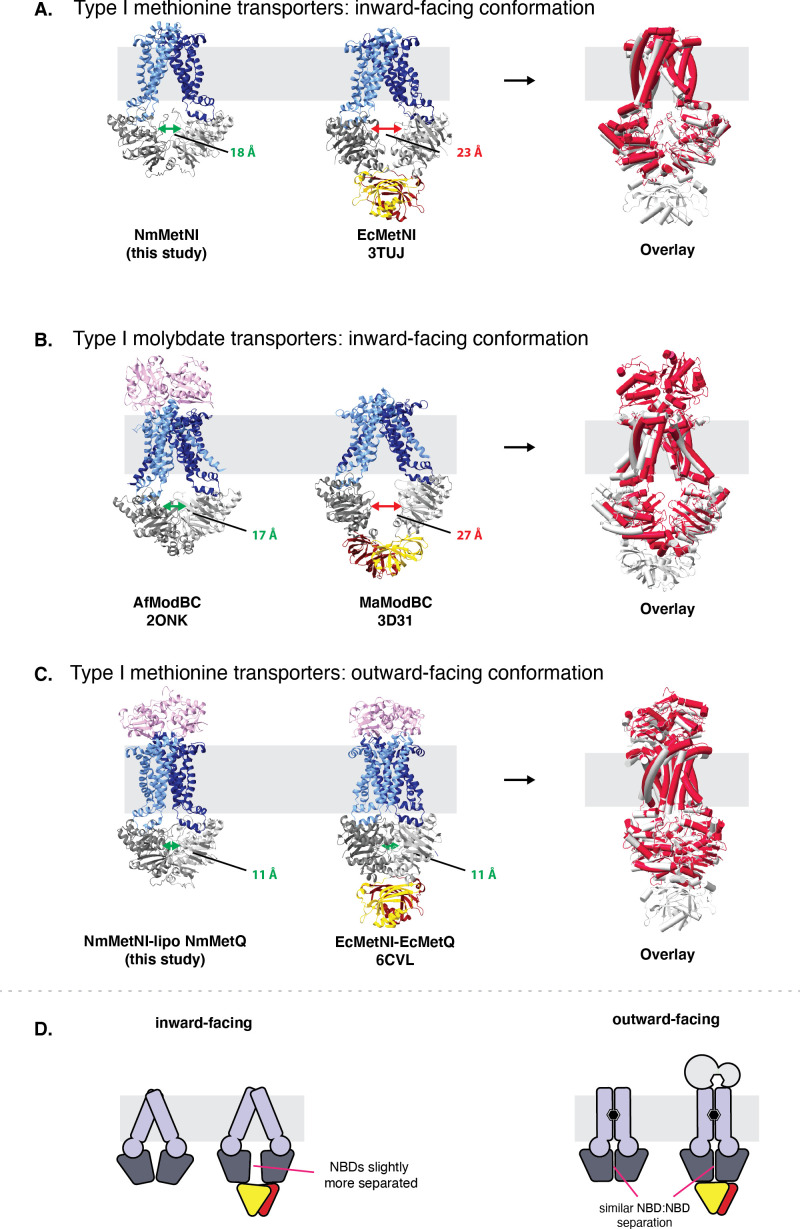
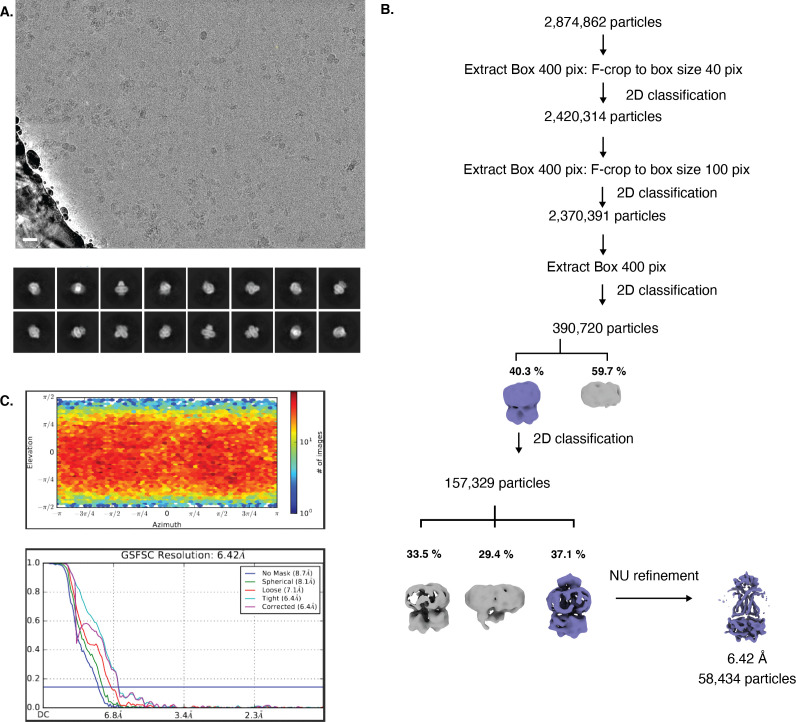
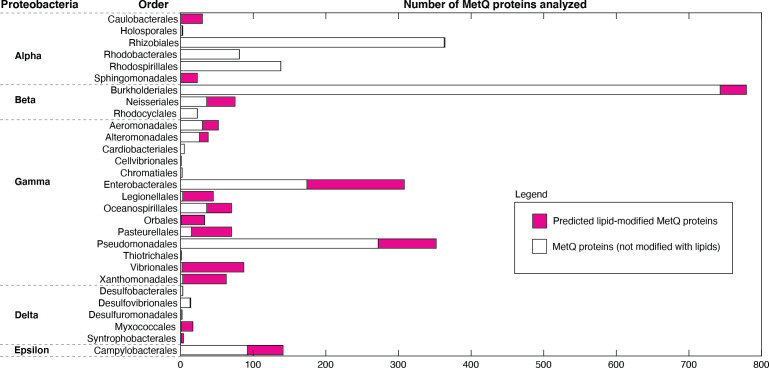
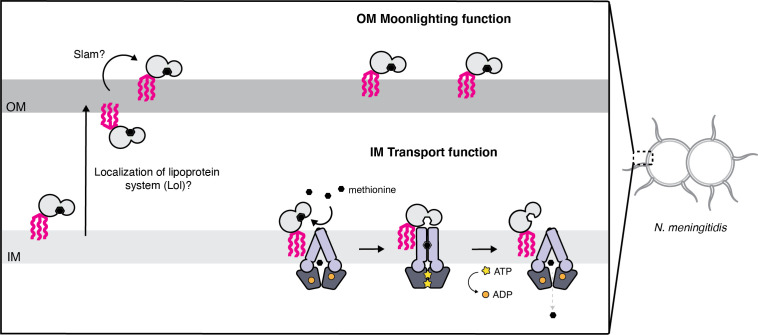
NmMetQ is a substrate-binding protein (SBP) from Neisseria meningitidis that has been identified as a surface-exposed candidate antigen for meningococcal vaccines. However, this location for NmMetQ challenges the prevailing view that SBPs in Gram-negative bacteria are localized to the periplasmic space to promote interaction with their cognate ABC transporter embedded in the bacterial inner membrane. To elucidate the roles of NmMetQ, we characterized NmMetQ with and without its cognate ABC transporter (NmMetNI). Here, we show that NmMetQ is a lipoprotein (lipo-NmMetQ) that binds multiple methionine analogs and stimulates the ATPase activity of NmMetNI. Using single-particle electron cryo-microscopy, we determined the structures of NmMetNI in the presence and absence of lipo-NmMetQ. Based on our data, we propose that NmMetQ tethers to membranes via a lipid anchor and has dual function and localization, playing a role in NmMetNI-mediated transport at the inner membrane and moonlighting on the bacterial surface.
Keywords: ABC transporters; E. coli; Neisseria meningitidis; biochemistry; chemical biology; fluorine solution nmr; lipoprotein; methionine importers; molecular biophysics; single particle cryoem; structural biology.
© 2021, Sharaf et al.
Conflict of interest statement
NS, MS, EK, JL, DV, AL, DR No competing interests declared
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
