

Accumulation of C-terminal cleaved tau is distinctly associated with cognitive deficits, synaptic plasticity impairment, and neurodegeneration in aged mice
- PMID: 34410588
- PMCID: PMC8810980
- DOI: 10.1007/s11357-021-00408-z
Accumulation of C-terminal cleaved tau is distinctly associated with cognitive deficits, synaptic plasticity impairment, and neurodegeneration in aged mice
Abstract
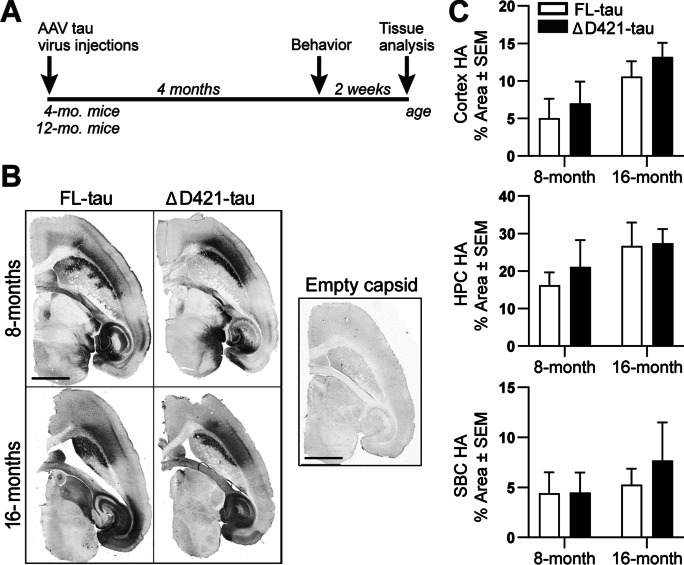
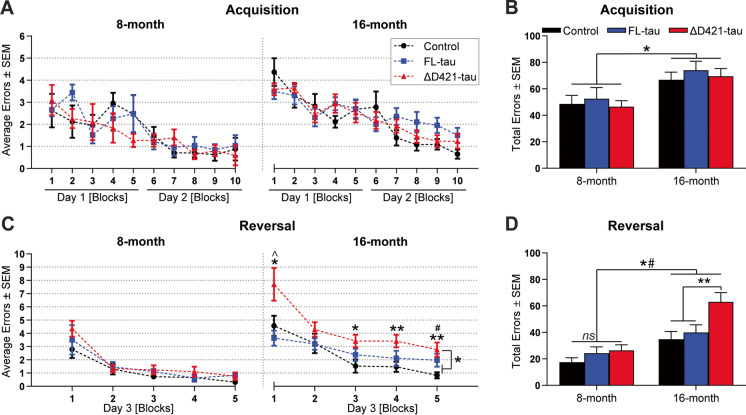
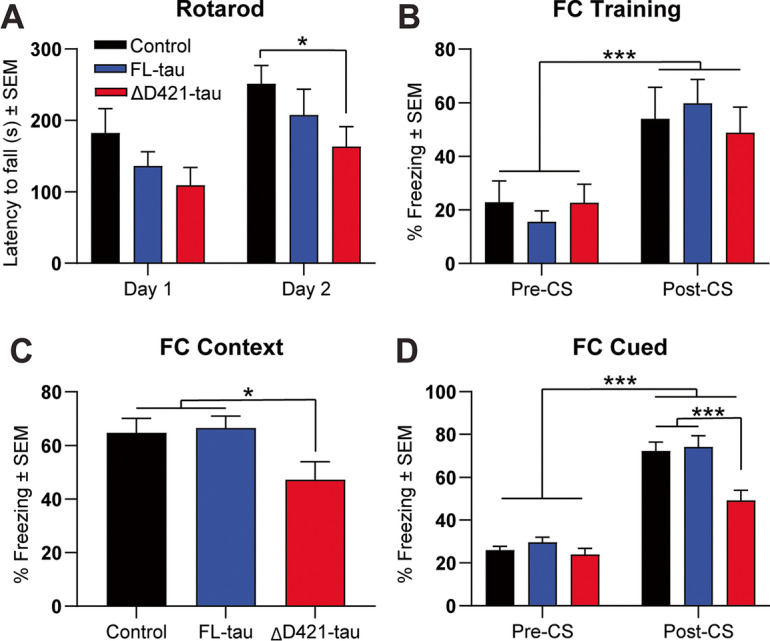
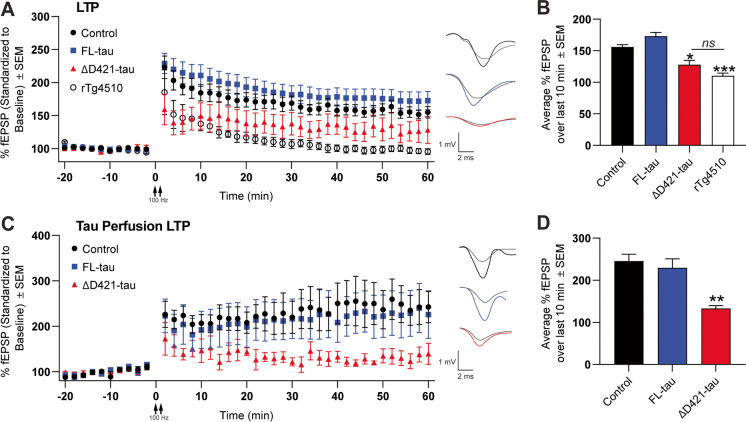
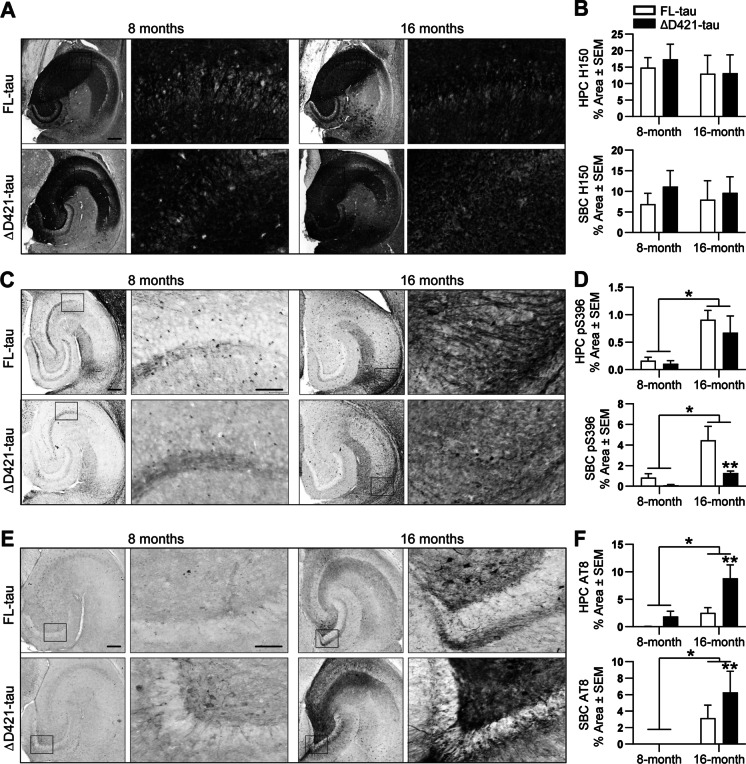
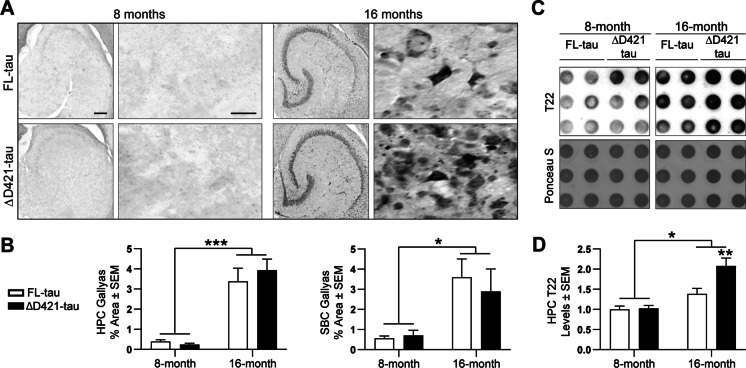
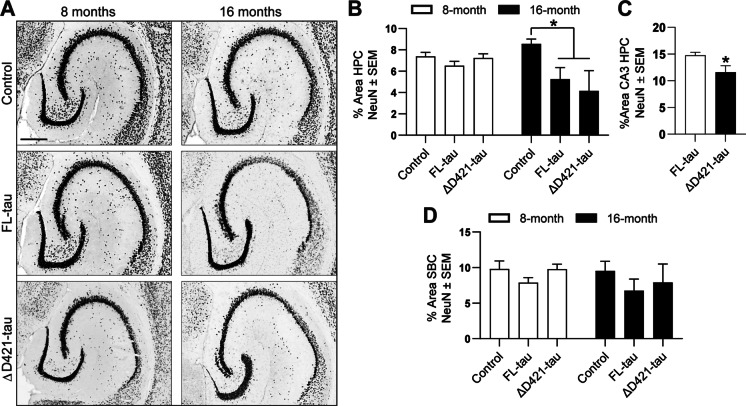
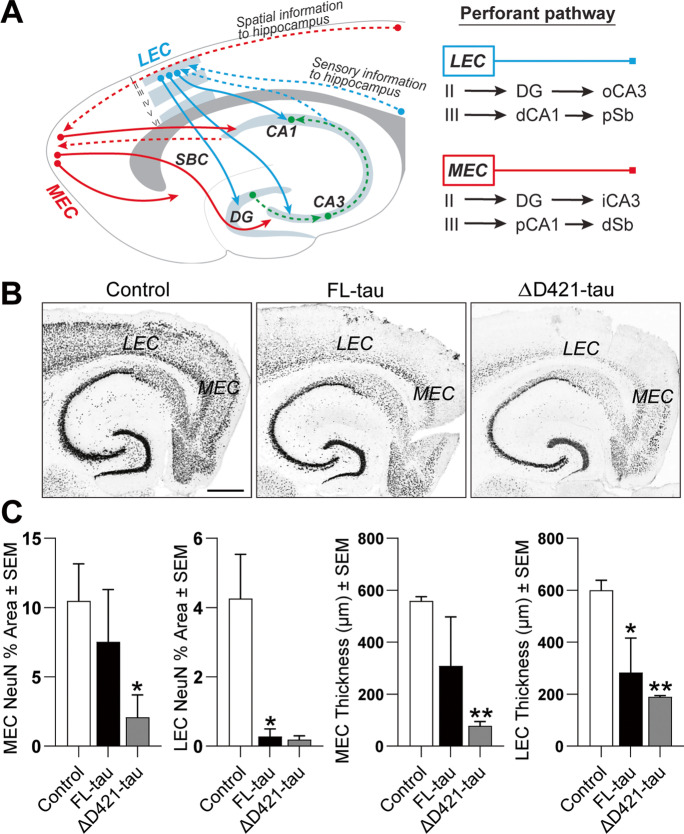
C-terminal cleaved tau at D421 (∆D421-tau) accumulates in the brains of Alzheimer's disease (AD) patients. However, it is unclear how tau truncation, an understudied tau post-translational modification, contributes to AD pathology and progression. Utilizing an adeno-associated virus (AAV) gene delivery-based approach, we overexpressed full-length tau (FL-tau) and ∆D421-tau in 4- and 12-month-old mice for 4 months to study the neuropathological impact of accumulation in young adult (8-month) and middle-aged (16-month) mice. Overall, we show that independent of the tau species, age was an important factor facilitating tau phosphorylation, oligomer formation, and deposition into silver-positive tangles. However, mice overexpressing ∆D421-tau exhibited a distinct phosphorylation profile to those overexpressing FL-tau and increased tau oligomerization in the middle-age group. Importantly, overexpression of ∆D421-tau, but not FL-tau in middle-aged mice, resulted in pronounced cognitive impairments and hippocampal long-term potentiation deficits. While both FL-tau and ∆D421-tau induced neuronal loss in mice with age, ∆D421-tau led to significant neuronal loss in the CA3 area of the hippocampus and medial entorhinal cortex compared to FL-tau. Based on our data, we conclude that age increases the susceptibility to neuronal degeneration associated with ΔD421-tau accumulation. Our findings suggest that ΔD421-tau accumulation contributes to synaptic plasticity and cognitive deficits, thus representing a potential target for tau-associated pathologies.
Keywords: Age; Cognition; Entorhinal cortex; Full-length tau; LTP; Neurodegeneration; Tauopathy; Truncated tau.
© 2021. American Aging Association.
Conflict of interest statement
The authors declare no competing interests.
Figures

Similar articles
-
Isoflurane mediated neuropathological and cognitive impairments in the triple transgenic Alzheimer's mouse model are associated with hippocampal synaptic deficits in an age-dependent manner.PLoS One. 2019 Oct 10;14(10):e0223509. doi: 10.1371/journal.pone.0223509. eCollection 2019. PLoS One. 2019. PMID: 31600350 Free PMC article.
-
STAT3 ameliorates cognitive deficits via regulation of NMDAR expression in an Alzheimer's disease animal model.Theranostics. 2021 Mar 13;11(11):5511-5524. doi: 10.7150/thno.56541. eCollection 2021. Theranostics. 2021. PMID: 33859760 Free PMC article.
-
Functional Alterations in the Olfactory Neuronal Circuit Occur before Hippocampal Plasticity Deficits in the P301S Mouse Model of Tauopathy: Implications for Early Diagnosis and Translational Research in Alzheimer's Disease.Int J Mol Sci. 2020 Jul 30;21(15):5431. doi: 10.3390/ijms21155431. Int J Mol Sci. 2020. PMID: 32751531 Free PMC article.
-
Regulatable transgenic mouse models of Alzheimer disease: onset, reversibility and spreading of Tau pathology.FEBS J. 2013 Sep;280(18):4371-81. doi: 10.1111/febs.12250. Epub 2013 Apr 22. FEBS J. 2013. PMID: 23517246 Review.
-
Neurodegeneration and plasticity.Int J Dev Neurosci. 2004 Nov;22(7):507-14. doi: 10.1016/j.ijdevneu.2004.07.007. Int J Dev Neurosci. 2004. PMID: 15465280 Review.
Cited by
-
Tau truncation in the pathogenesis of Alzheimer's disease: a narrative review.Neural Regen Res. 2024 Jun 1;19(6):1221-1232. doi: 10.4103/1673-5374.385853. Epub 2023 Sep 22. Neural Regen Res. 2024. PMID: 37905868 Free PMC article.
-
Pathological Impact of Tau Proteolytical Process on Neuronal and Mitochondrial Function: a Crucial Role in Alzheimer's Disease.Mol Neurobiol. 2023 Oct;60(10):5691-5707. doi: 10.1007/s12035-023-03434-4. Epub 2023 Jun 19. Mol Neurobiol. 2023. PMID: 37332018 Review.
-
Adeno-Associated Viral Gene Delivery of Wild-Type Human Tau Induces Progressive Hyperphosphorylation and Neuronal Cell Death in the Hippocampi of Middle-Aged Rats.Cells. 2025 Aug 11;14(16):1238. doi: 10.3390/cells14161238. Cells. 2025. PMID: 40862717 Free PMC article.
-
Tau Post-Translational Modifications: Potentiators of Selective Vulnerability in Sporadic Alzheimer's Disease.Biology (Basel). 2021 Oct 15;10(10):1047. doi: 10.3390/biology10101047. Biology (Basel). 2021. PMID: 34681146 Free PMC article. Review.
-
Depletion of TDP-43 exacerbates tauopathy-dependent brain atrophy by sensitizing vulnerable neurons to caspase 3-mediated endoproteolysis of tau in a mouse model of Multiple Etiology Dementia.bioRxiv [Preprint]. 2024 Jun 29:2024.06.26.600814. doi: 10.1101/2024.06.26.600814. bioRxiv. 2024. PMID: 38979270 Free PMC article. Preprint.
References
-
- Abisambra J, Jinwal UK, Miyata Y, Rogers J, Blair L, Li X, Seguin SP, Wang L, Jin Y, Bacon J, Brady S, Cockman M, Guidi C, Zhang J, Koren J, Young ZT, Atkins CA, Zhang B, Lawson LY, Weeber EJ, Brodsky JL, Gestwicki JE, Dickey CA. Allosteric heat shock protein 70 inhibitors rapidly rescue synaptic plasticity deficits by reducing aberrant tau. Biol Psychiatry. 2013. 10.1016/j.biopsych.2013.02.027 - PMC - PubMed
-
- Abraha A, Ghoshal N, Gamblin TC, Cryns V, Berry RW, Kuret J, Binder LI. C-terminal inhibition of tau assembly in vitro and in Alzheimer’s disease. J Cell Sci. 2000. 10.1242/jcs.113.21.3737 - PubMed
-
- Ahmed Z, Cooper J, Murray TK, Garn K, McNaughton E, Clarke H, Parhizkar S, Ward MA, Cavallini A, Jackson S, Bose S, Clavaguera F, Tolnay M, Lavenir I, Goedert M, Hutton ML, O’Neill MJ. A novel in vivo model of tau propagation with rapid and progressive neurofibrillary tangle pathology: The pattern of spread is determined by connectivity, not proximity. Acta Neuropathol. 2014 doi: 10.1007/s00401-014-1254-6. - DOI - PMC - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
