

IL-33 activates mTORC1 and modulates glycolytic metabolism in CD8+ T cells
- PMID: 34411293
- PMCID: PMC9112898
- DOI: 10.1111/imm.13404
IL-33 activates mTORC1 and modulates glycolytic metabolism in CD8+ T cells
Abstract
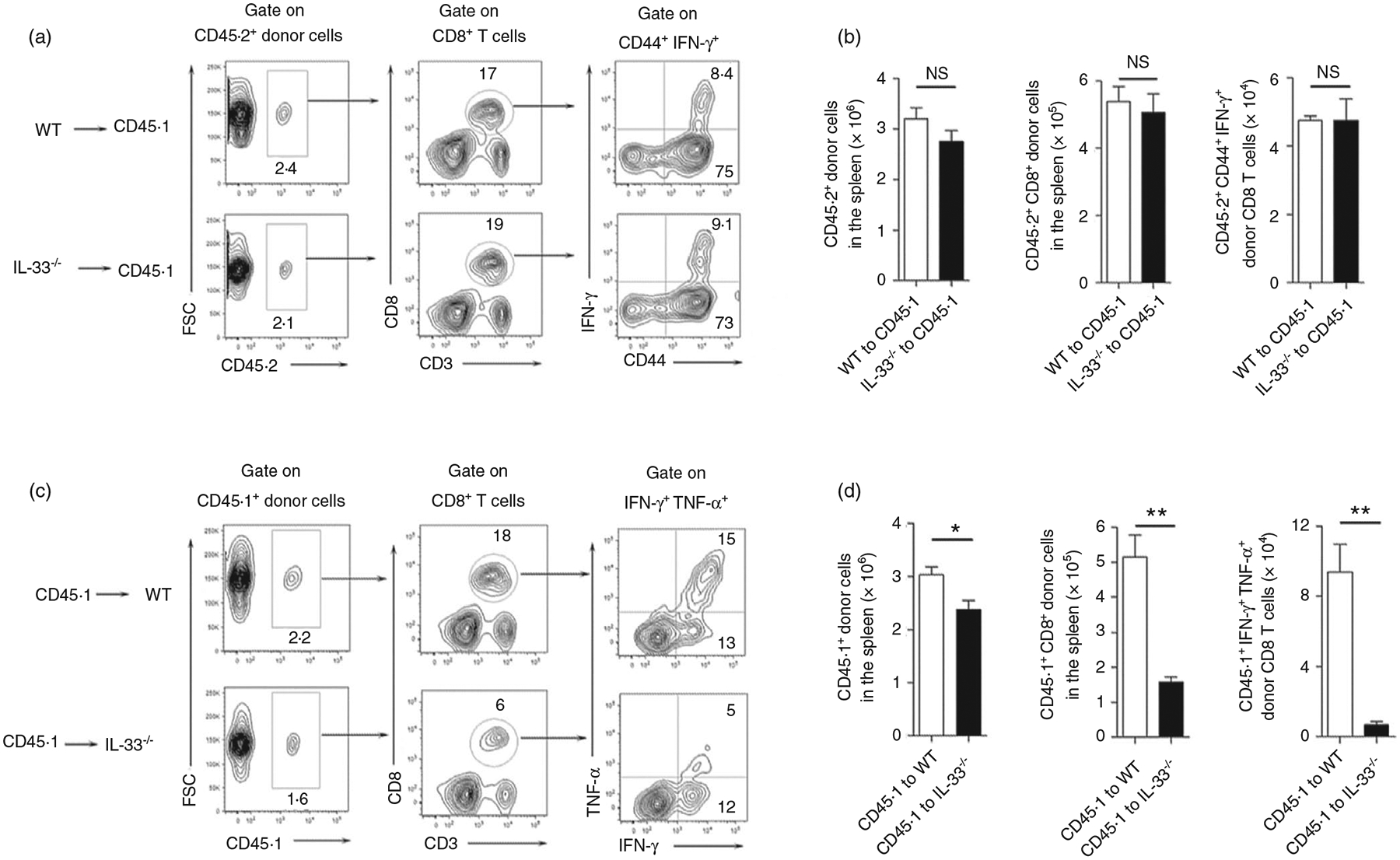
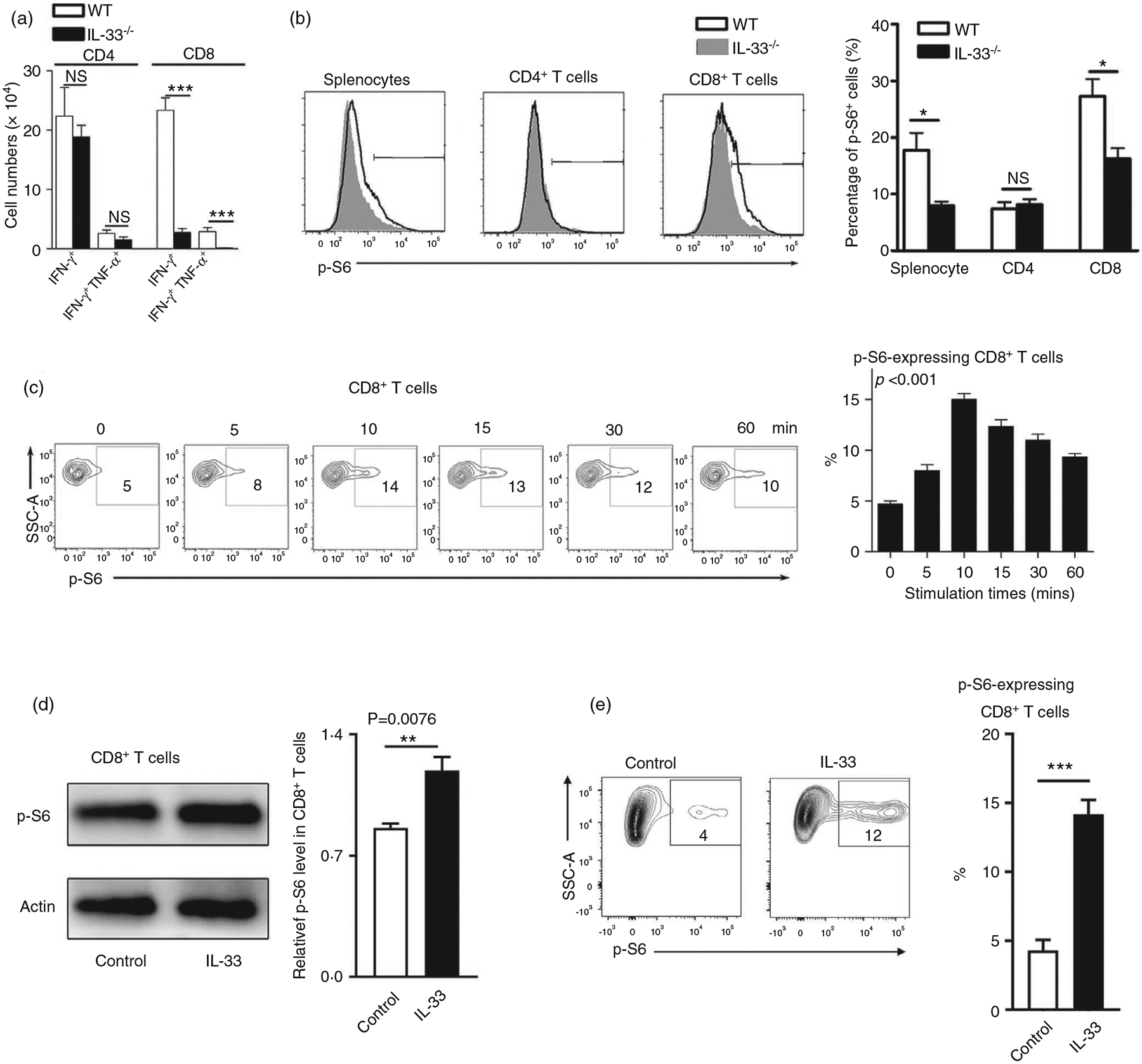
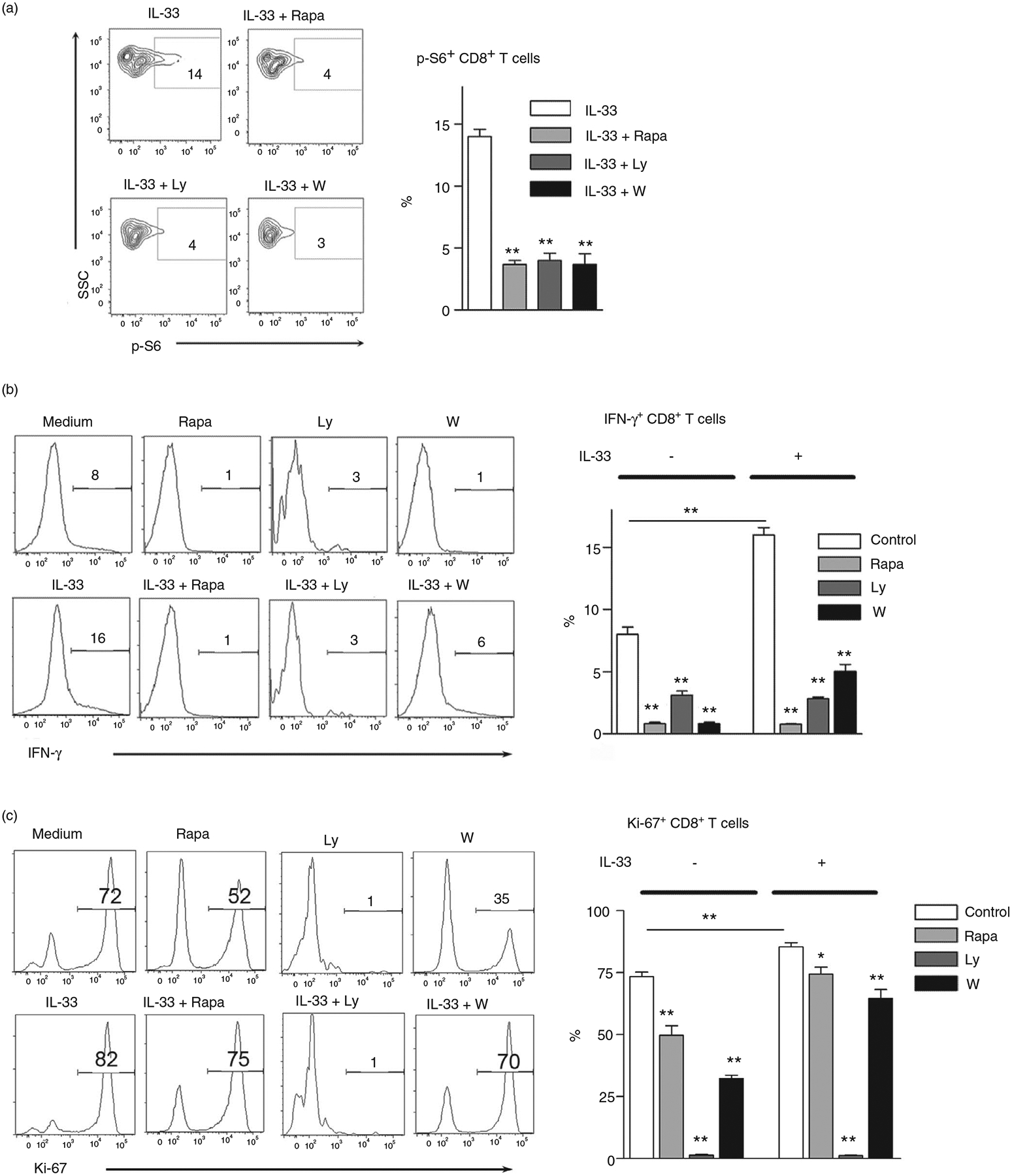
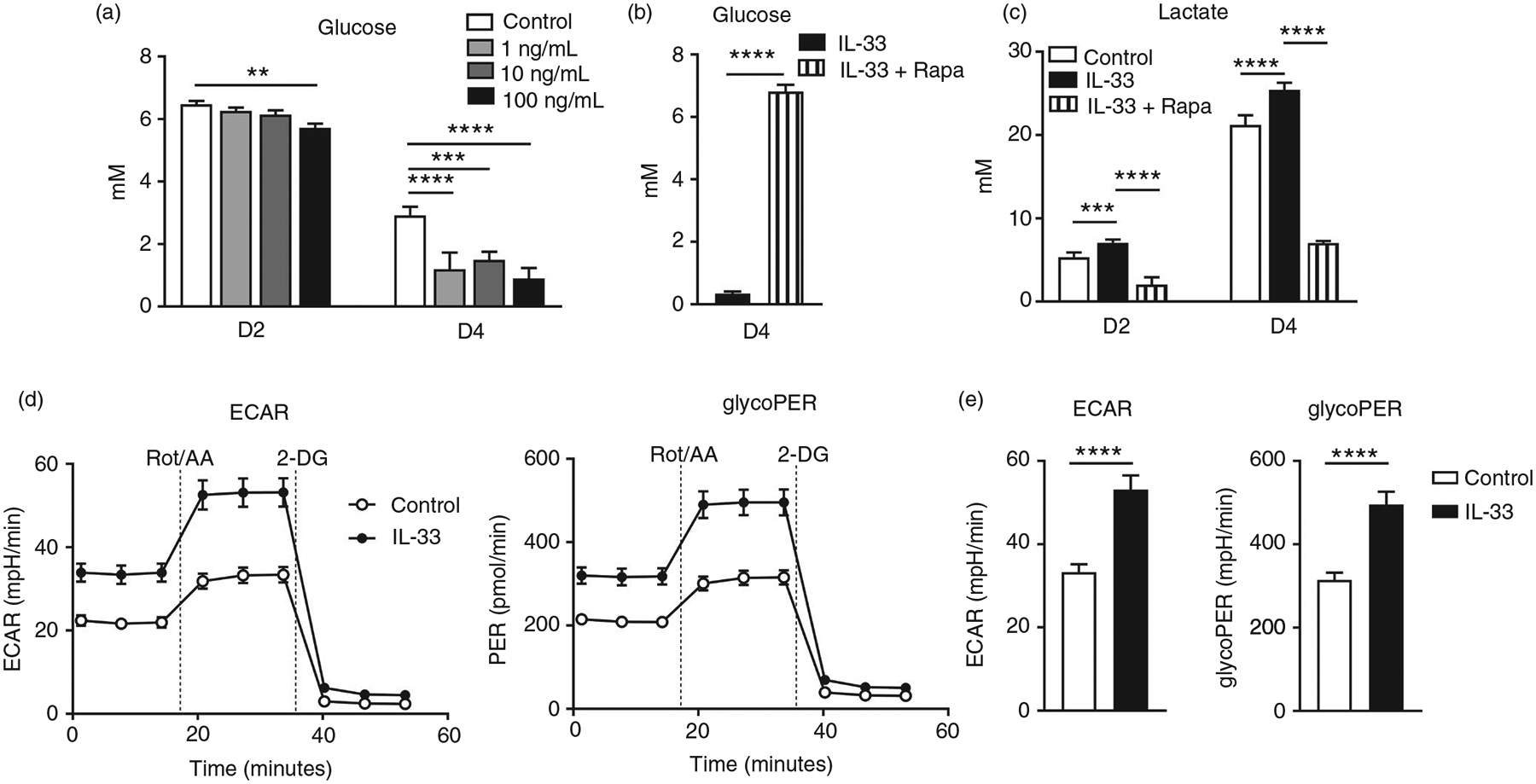
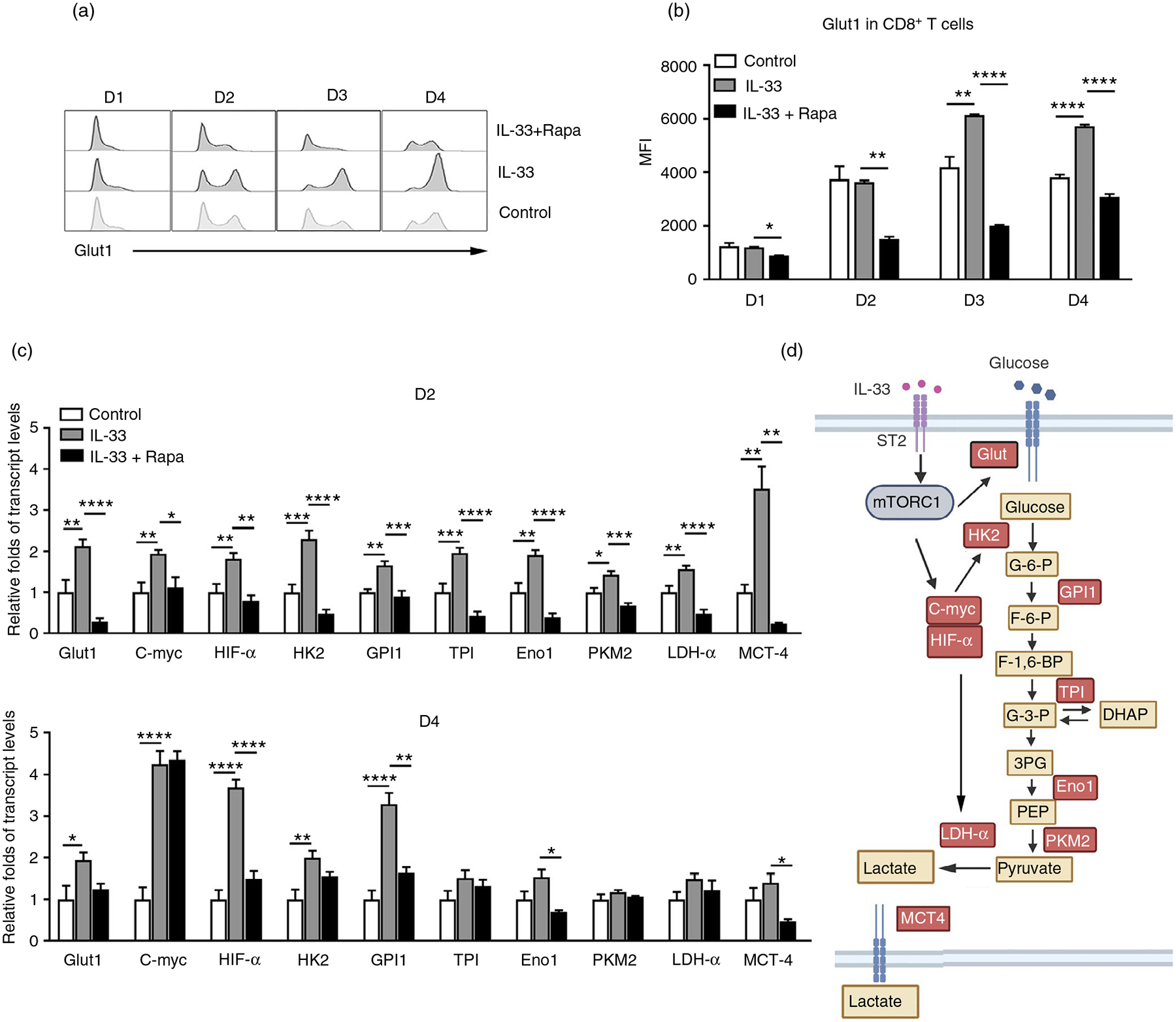
Interleukin (IL)-33, a member in the IL-1 family, plays a central role in innate and adaptive immunity; however, how IL-33 mediates cytotoxic T-cell regulation and the downstream signals remain elusive. In this study, we found increased mouse IL-33 expression in CD8+ T cells following cell activation via anti-CD3/CD28 stimulation in vitro or lymphocytic choriomeningitis virus (LCMV) infection in vivo. Our cell adoptive transfer experiment demonstrated that extracellular, but not nuclear, IL-33 contributed to the activation and proliferation of CD8+ , but not CD4+ T effector cells in LCMV infection. Importantly, IL-33 induced mTORC1 activation in CD8+ T cells as evidenced by increased phosphorylated S6 ribosomal protein (p-S6) levels both in vitro and in vivo. Meanwhile, this IL-33-induced CD8+ T-cell activation was suppressed by mTORC1 inhibitors. Furthermore, IL-33 elevated glucose uptake and lactate production in CD8+ T cells in both dose- and time-dependent manners. The results of glycolytic rate assay demonstrated the increased glycolytic capacity of IL-33-treated CD8+ T cells compared with that of control cells. Our mechanistic study further revealed the capacity of IL-33 in promoting the expression of glucose transporter 1 (Glut1) and glycolytic enzymes via mTORC1, leading to accelerated aerobic glucose metabolism Warburg effect and increased effector T-cell activation. Together, our data provide new insights into IL-33-mediated regulation of CD8+ T cells, which might be beneficial for therapeutic strategies of inflammatory and infectious diseases in the future.
Keywords: CD8; Glut1; IL-33; T cells; glycolytic metabolism; mTORC1.
© 2021 John Wiley & Sons Ltd.
Conflict of interest statement
CONFLICT OF INTEREST
The authors have no financial conflicts of interest.
Figures
References
-
- Schmitz J, Owyang A, Oldham E, Song Y, Murphy E, McClanahan TK, et al. IL-33, an interleukin-1-like cytokine that signals via the IL-1 receptor-related protein ST2 and induces T helper type 2-associated cytokines. Immunity. 2005;23:479–90. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
