

Targeting Cpt1a-Bcl-2 interaction modulates apoptosis resistance and fibrotic remodeling
- PMID: 34413485
- PMCID: PMC8738732
- DOI: 10.1038/s41418-021-00840-w
Targeting Cpt1a-Bcl-2 interaction modulates apoptosis resistance and fibrotic remodeling
Abstract
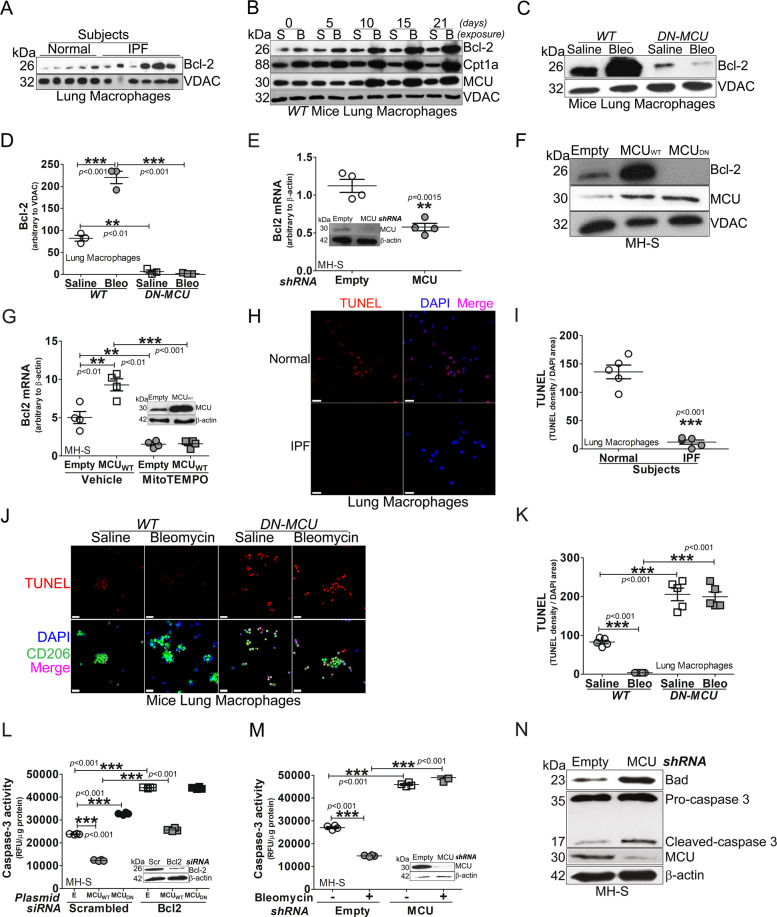
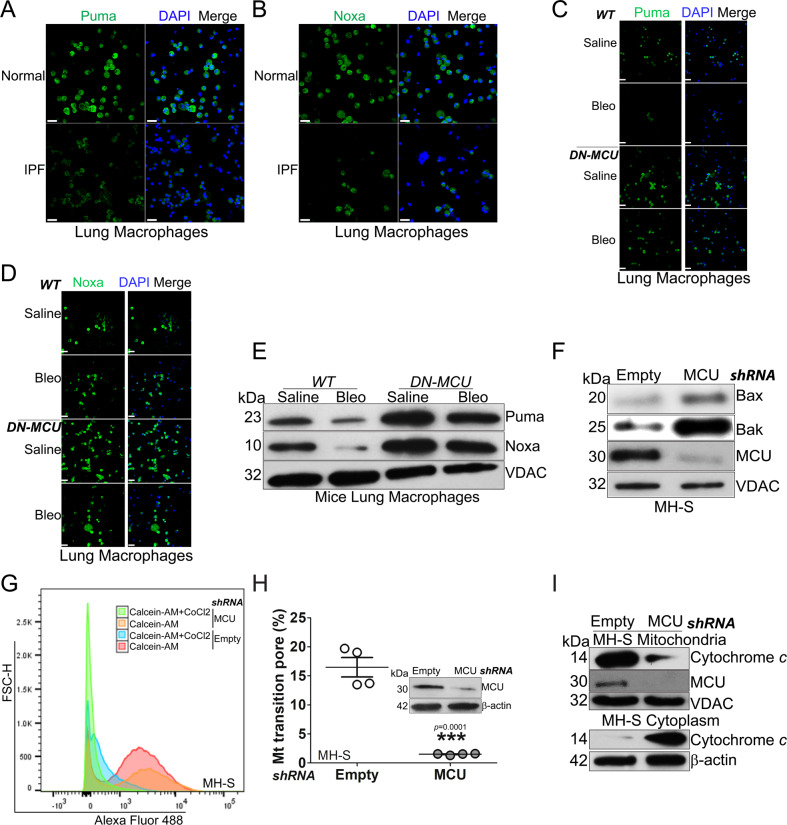
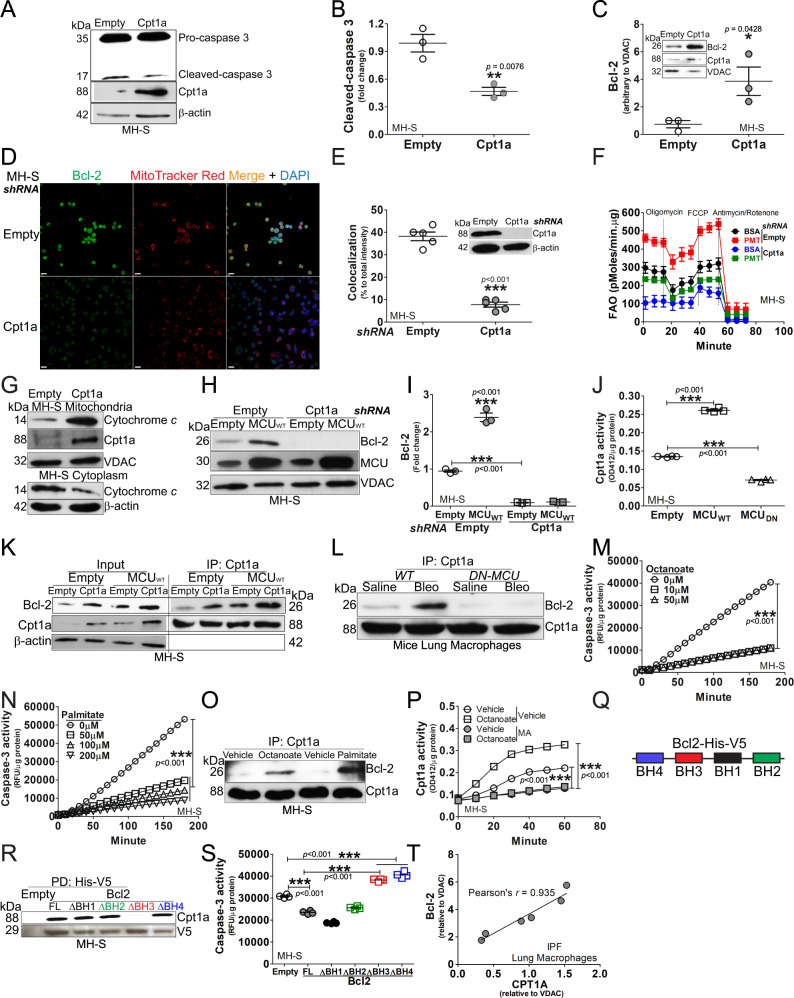
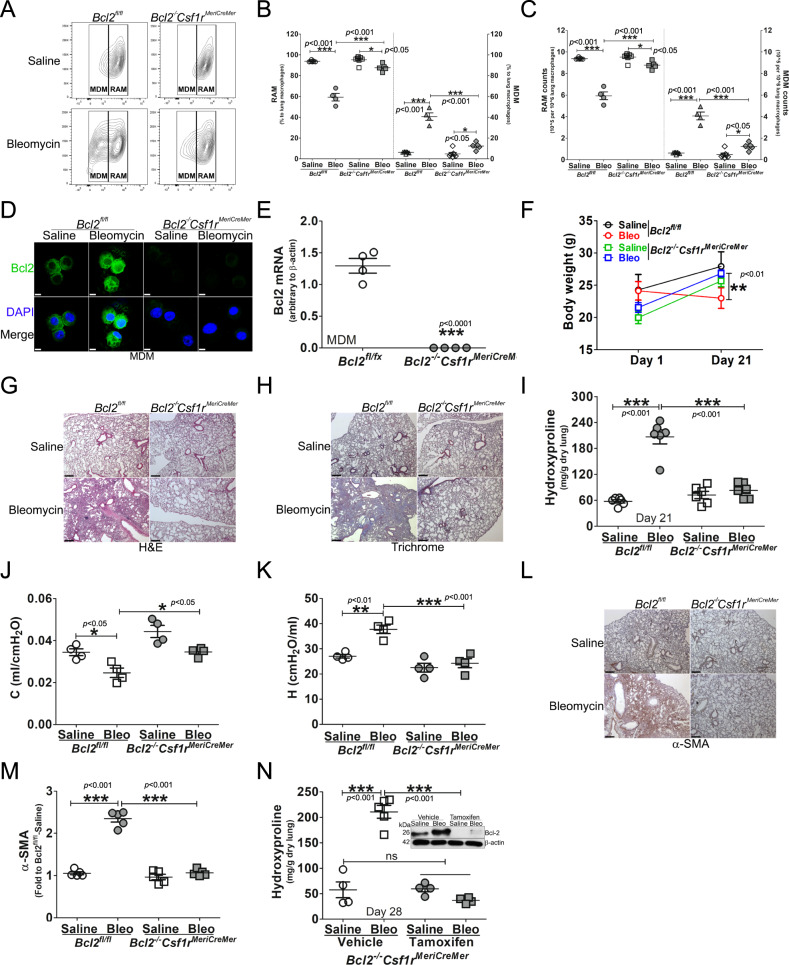
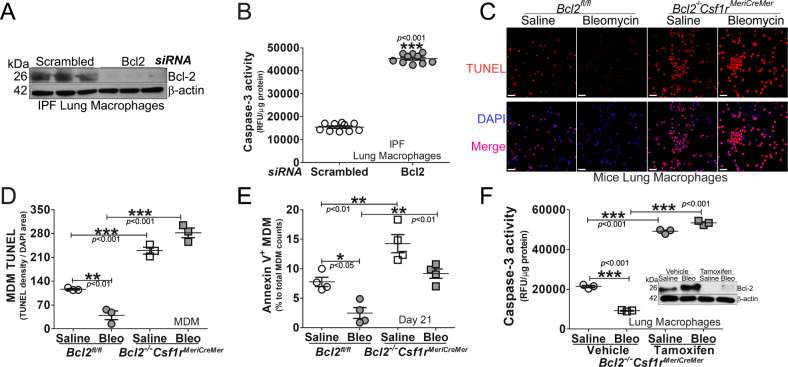
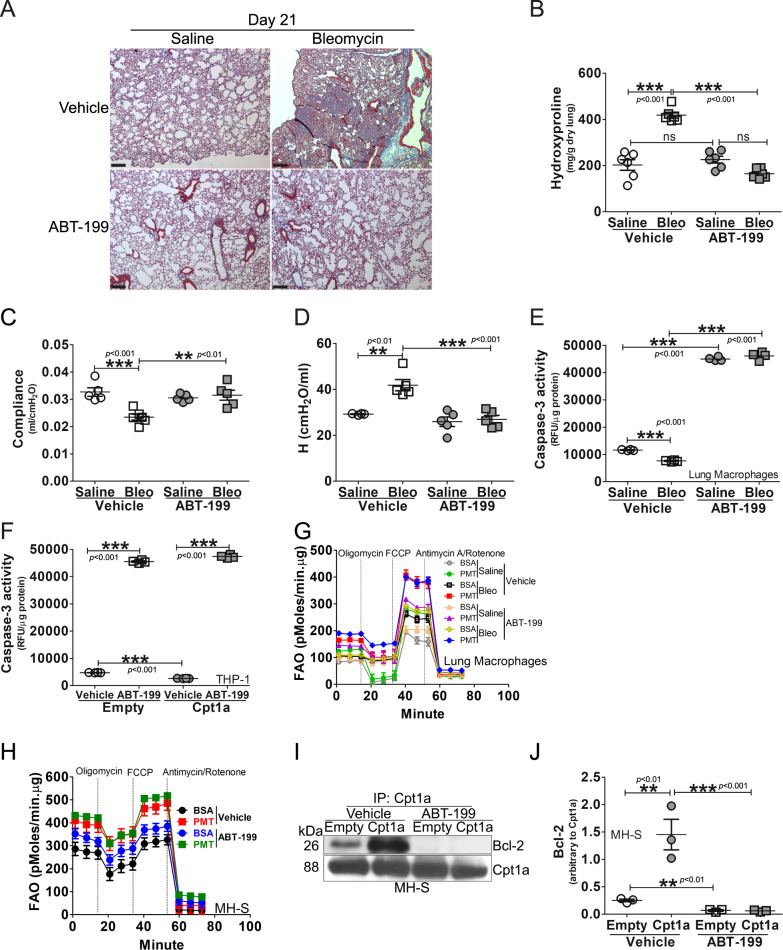
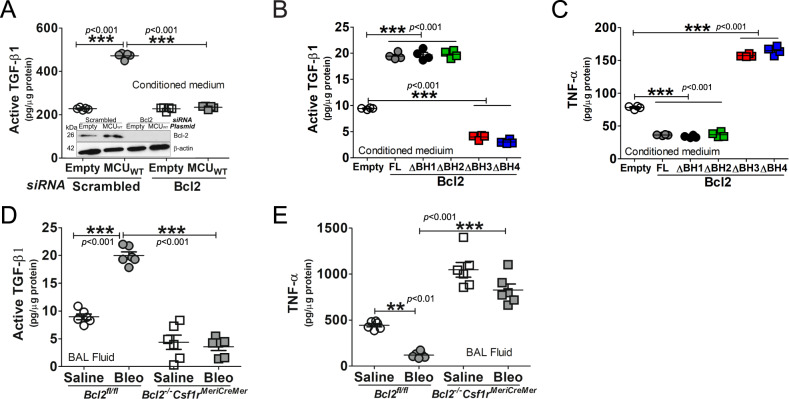
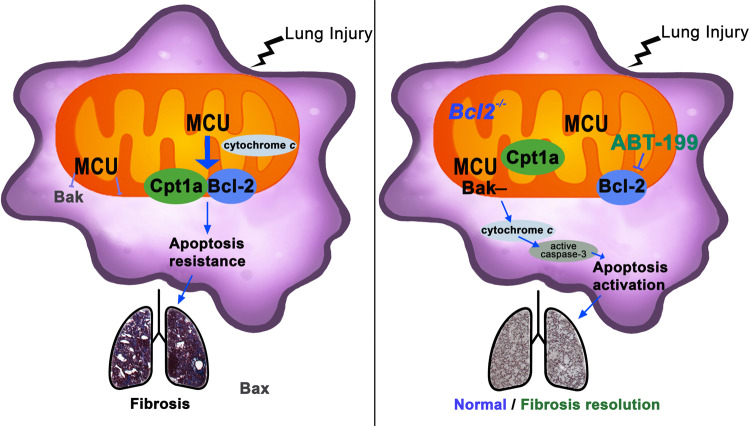
The mitochondrial calcium uniporter (MCU) regulates metabolic reprogramming in lung macrophages and the progression of pulmonary fibrosis. Fibrosis progression is associated with apoptosis resistance in lung macrophages; however, the mechanism(s) by which apoptosis resistance occurs is poorly understood. Here, we found a marked increase in mitochondrial B-cell lymphoma-2 (Bcl-2) in lung macrophages from subjects with idiopathic pulmonary fibrosis (IPF). Similar findings were seen in bleomycin-injured wild-type (WT) mice, whereas Bcl-2 was markedly decreased in mice expressing a dominant-negative mitochondrial calcium uniporter (DN-MCU). Carnitine palmitoyltransferase 1a (Cpt1a), the rate-limiting enzyme for fatty acid β-oxidation, directly interacted with Bcl-2 by binding to its BH3 domain, which anchored Bcl-2 in the mitochondria to attenuate apoptosis. This interaction was dependent on Cpt1a activity. Lung macrophages from IPF subjects had a direct correlation between CPT1A and Bcl-2, whereas the absence of binding induced apoptosis. The deletion of Bcl-2 in macrophages protected mice from developing pulmonary fibrosis. Moreover, mice had resolution when Bcl-2 was deleted or was inhibited with ABT-199 after fibrosis was established. These observations implicate an interplay between macrophage fatty acid β-oxidation, apoptosis resistance, and dysregulated fibrotic remodeling.
© 2021. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Lederer DJ, Martinez FJ. Idiopathic pulmonary fibrosis. N. Engl J Med. 2018;379:797–8. - PubMed
-
- King TE, Jr., Bradford WZ, Castro-Bernardini S, Fagan EA, Glaspole I, Glassberg MK, et al. A phase 3 trial of pirfenidone in patients with idiopathic pulmonary fibrosis. N. Engl J Med. 2014;370:2083–92. - PubMed
-
- Richeldi L, du Bois RM, Raghu G, Azuma A, Brown KK, Costabel U, et al. Efficacy and safety of nintedanib in idiopathic pulmonary fibrosis. N. Engl J Med. 2014;370:2071–82. - PubMed
-
- Docherty NG, O’Sullivan OE, Healy DA, Fitzpatrick JM, Watson RW. Evidence that inhibition of tubular cell apoptosis protects against renal damage and development of fibrosis following ureteric obstruction. Am J Physiol Ren Physiol. 2006;290:F4–13. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
