

What defines the maternal transcriptome?
- PMID: 34415300
- PMCID: PMC8589422
- DOI: 10.1042/BST20201125
What defines the maternal transcriptome?
Abstract
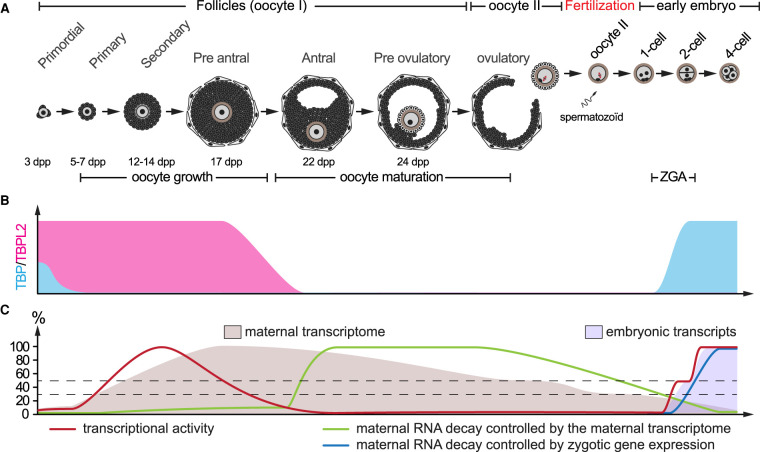
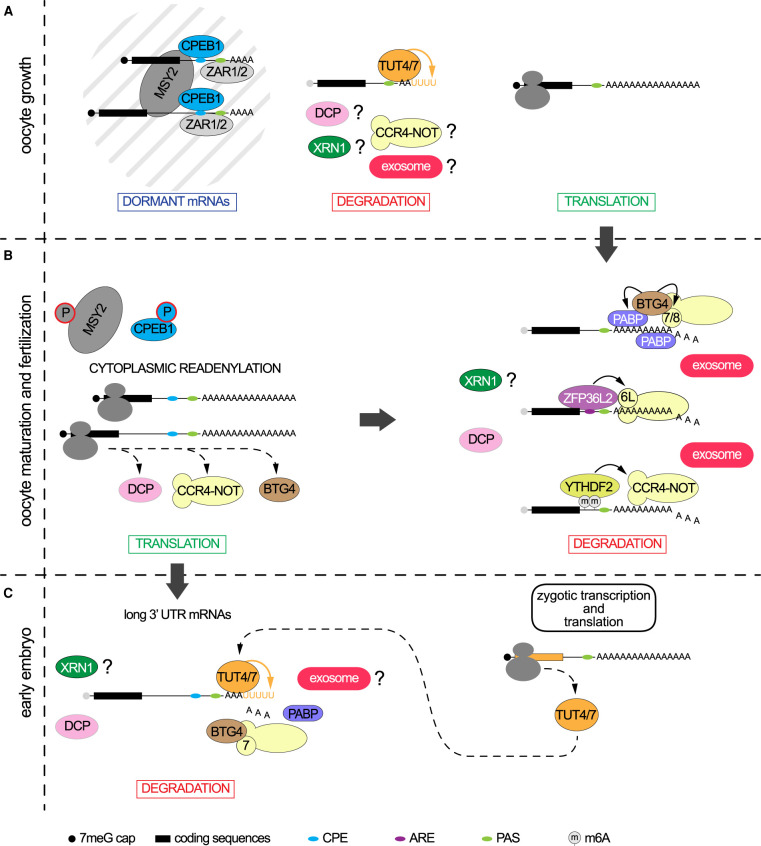
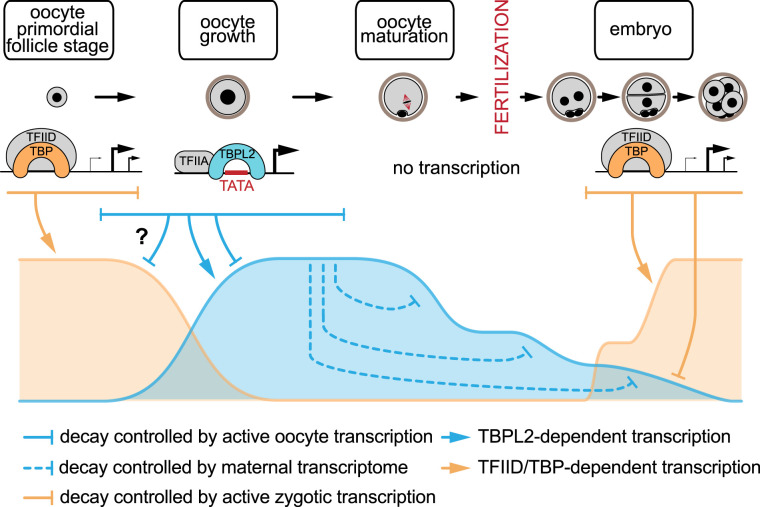
In somatic cells, RNA polymerase II (Pol II) transcription initiation starts by the binding of the general transcription factor TFIID, containing the TATA-binding protein (TBP) and 13 TBP-associated factors (TAFs), to core promoters. However, in growing oocytes active Pol II transcription is TFIID/TBP-independent, as during oocyte growth TBP is replaced by its vertebrate-specific paralog TBPL2. TBPL2 does not interact with TAFs, but stably associates with TFIIA. The maternal transcriptome is the population of mRNAs produced and stored in the cytoplasm of growing oocytes. After fertilization, maternal mRNAs are inherited by the zygote from the oocyte. As transcription becomes silent after oocyte growth, these mRNAs are the sole source for active protein translation. They will participate to complete the protein pool required for oocyte terminal differentiation, fertilization and initiation of early development, until reactivation of transcription in the embryo, called zygotic genome activation (ZGA). All these events are controlled by an important reshaping of the maternal transcriptome. This procedure combines cytoplasmic readenylation of stored transcripts, allowing their translation, and different waves of mRNA degradation by deadenylation coupled to decapping, to eliminate transcripts coding for proteins that are no longer required. The reshaping ends after ZGA with an almost total clearance of the maternal transcripts. In the past, the murine maternal transcriptome has received little attention but recent progresses have brought new insights into the regulation of maternal mRNA dynamics in the mouse. This review will address past and recent data on the mechanisms associated with maternal transcriptome dynamic in the mouse.
Keywords: RNA decay; RNA polymerase II transcription; TBPL2; early embryo; mouse oocytes; readenylation.
© 2021 The Author(s).
Conflict of interest statement
The authors declare that there are no competing interests associated with the manuscript.
Figures
References
-
- Kidder, G.M. and Pedersen, R.A. (1982) Turnover of embryonic messenger RNA in preimplantation mouse embryos. J. Embryol. Exp. Morph. 67, 37–49 PMID: - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous