

Therapeutic alphavirus cross-reactive E1 human antibodies inhibit viral egress
- PMID: 34416147
- PMCID: PMC8418820
- DOI: 10.1016/j.cell.2021.07.033
Therapeutic alphavirus cross-reactive E1 human antibodies inhibit viral egress
Abstract
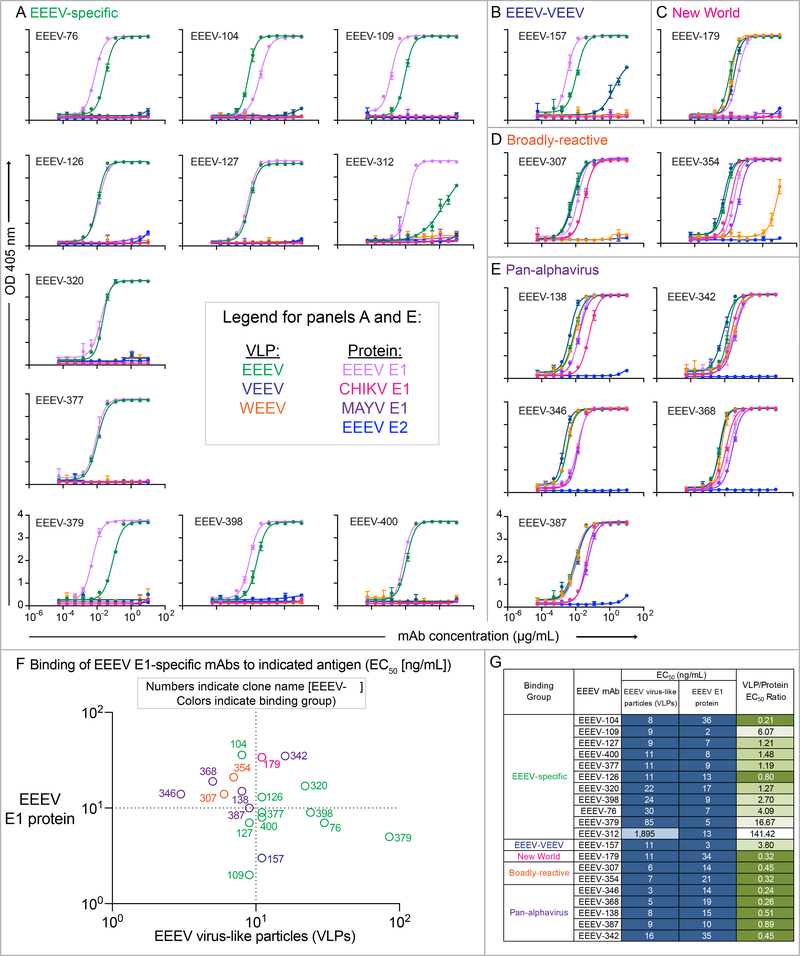
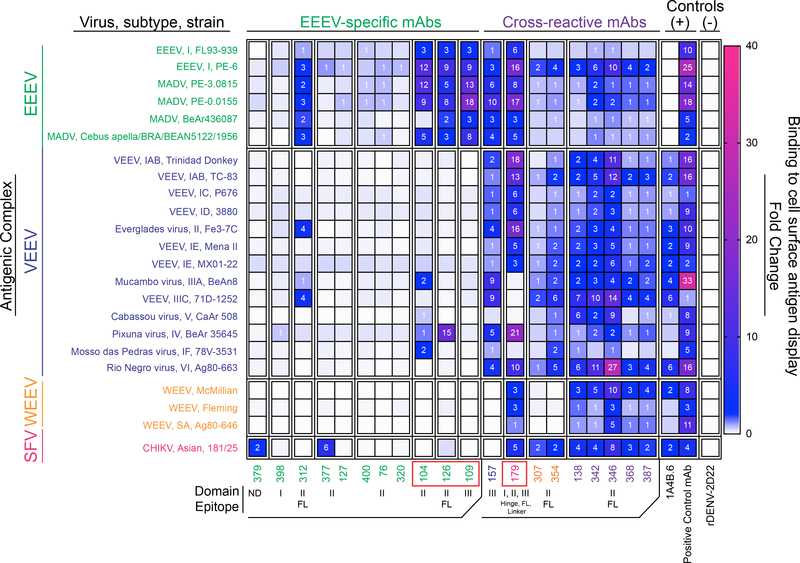
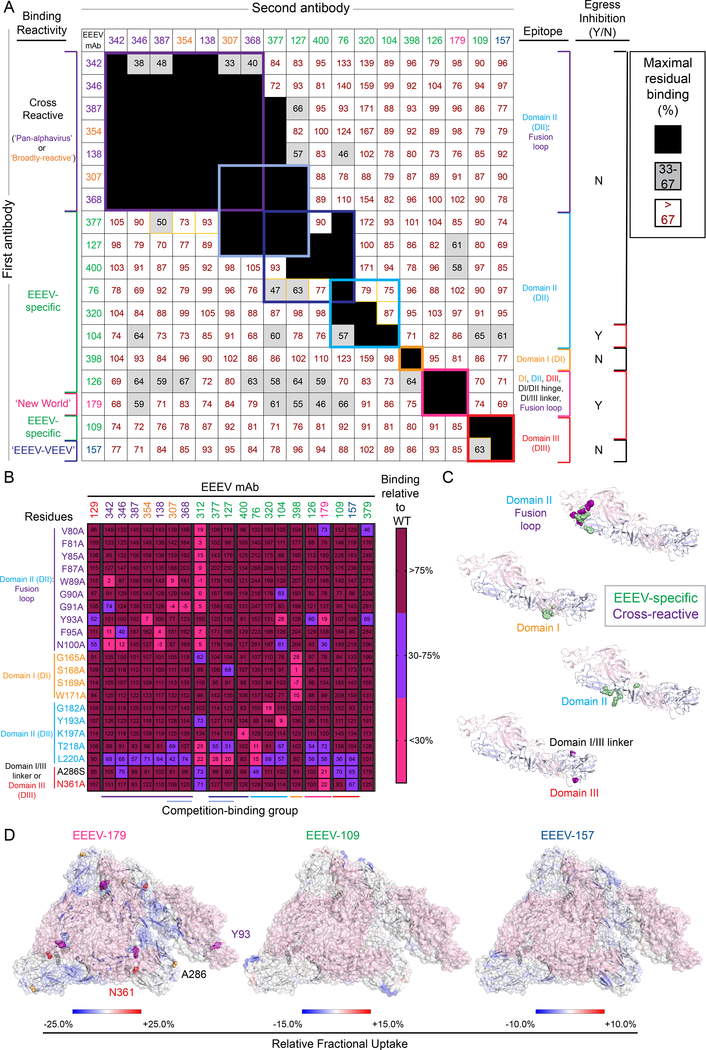
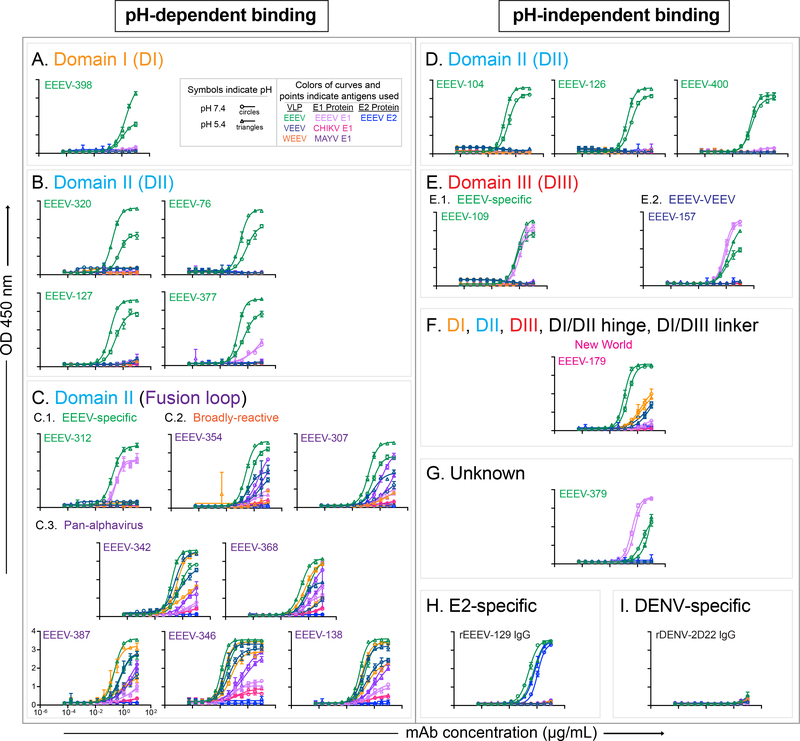
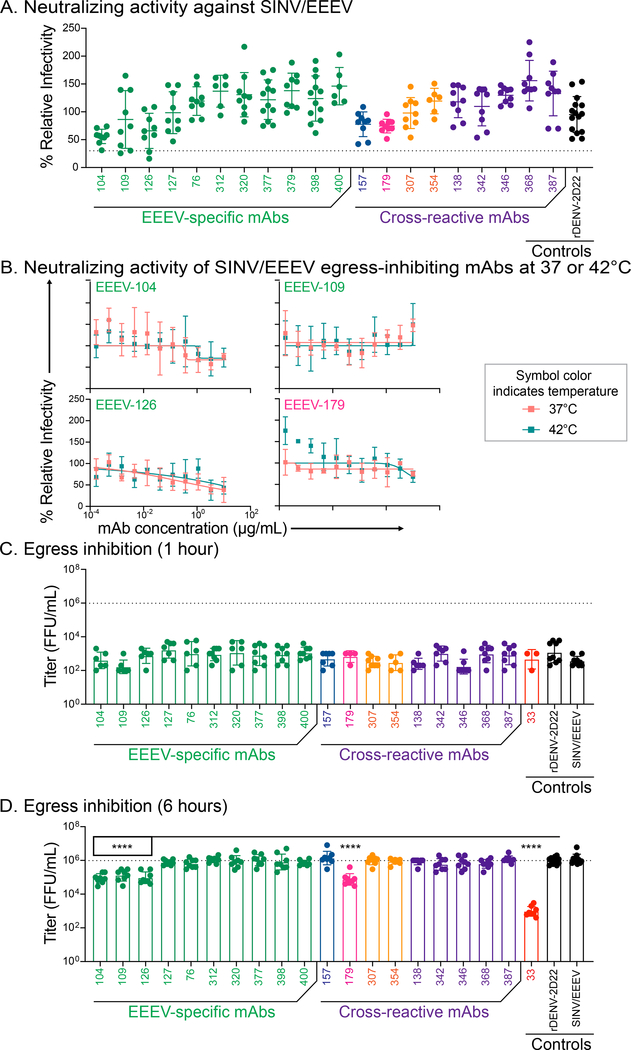
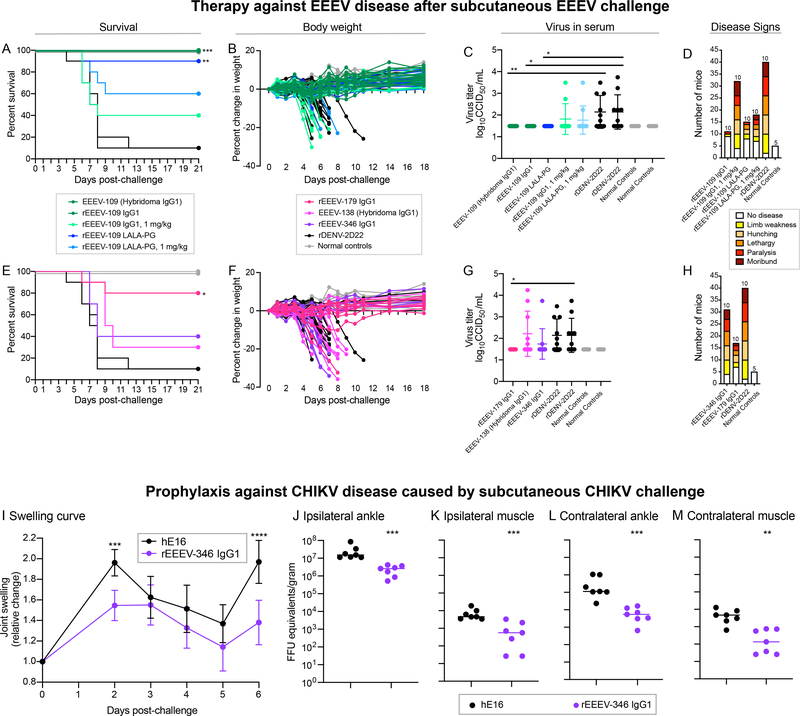
Alphaviruses cause severe arthritogenic or encephalitic disease. The E1 structural glycoprotein is highly conserved in these viruses and mediates viral fusion with host cells. However, the role of antibody responses to the E1 protein in immunity is poorly understood. We isolated E1-specific human monoclonal antibodies (mAbs) with diverse patterns of recognition for alphaviruses (ranging from Eastern equine encephalitis virus [EEEV]-specific to alphavirus cross-reactive) from survivors of natural EEEV infection. Antibody binding patterns and epitope mapping experiments identified differences in E1 reactivity based on exposure of epitopes on the glycoprotein through pH-dependent mechanisms or presentation on the cell surface prior to virus egress. Therapeutic efficacy in vivo of these mAbs corresponded with potency of virus egress inhibition in vitro and did not require Fc-mediated effector functions for treatment against subcutaneous EEEV challenge. These studies reveal the molecular basis for broad and protective antibody responses to alphavirus E1 proteins.
Keywords: alphavirus; cross-reactive antibodies; human monoclonal antibodies; non-neutralizing antibodies; post-exposure prophylaxis.
Copyright © 2021 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests J.E.C. has served as a consultant for Luna Biologics, is a member of the Scientific Advisory Board of Meissa Vaccines, and is Founder of IDBiologics. The Crowe laboratory at Vanderbilt University Medical Center has received unrelated sponsored research agreements from IDBiologics and AstraZeneca. M.S.D. is a consultant for Inbios, Vir Biotechnology, NGM Biopharmaceuticals, and Carnival Corporation and on the Scientific Advisory Boards of Moderna and Immunome. The Diamond laboratory at Washington University School of Medicine has received sponsored research agreements from Emergent BioSolutions, Moderna, and Vir Biotechnology. All other authors report no conflicts. M.E.F., E.D., and B.J.D. are employees of Integral Molecular and B.J.D. is a shareholder. G.A. is a founder and the Chief Scientific Officer (Consulting) of Seromyx.
Figures
References
-
- Aguilar-Luis MA, Del Valle-Mendoza J, Silva-Caso W, Gil-Ramirez T, Levy-Blitchtein S,Bazan-Mayra J, Zavaleta-Gavidia V, Cornejo-Pacherres D, Palomares-Reyes C, and Del Valle LJ (2020). An emerging public health threat: Mayaro virus increases its distribution in Peru. Int J Infect Dis 92, 253–258. - PubMed
-
- Armstrong PM, and Andreadis TG (2013). Eastern equine encephalitis virus--old enemy, new threat. N Engl J Med 368, 1670–1673. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
