

Subsynaptic positioning of AMPARs by LRRTM2 controls synaptic strength
- PMID: 34417170
- PMCID: PMC8378824
- DOI: 10.1126/sciadv.abf3126
Subsynaptic positioning of AMPARs by LRRTM2 controls synaptic strength
Abstract
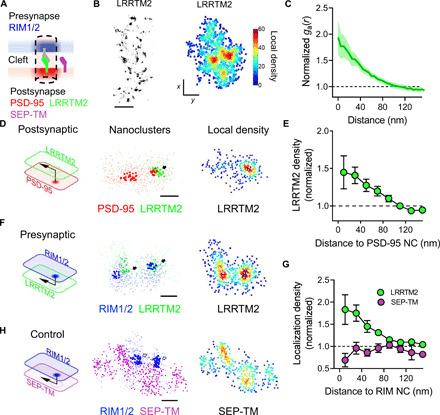
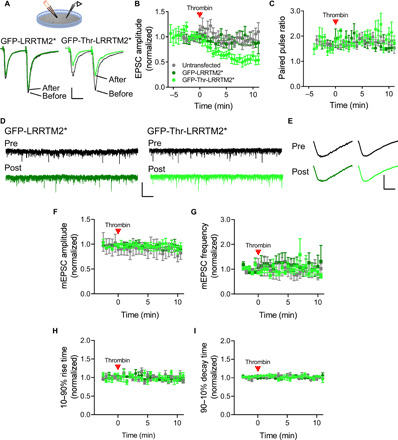
Recent evidence suggests that nano-organization of proteins within synapses may control the strength of communication between neurons in the brain. The unique subsynaptic distribution of glutamate receptors, which cluster in nanoalignment with presynaptic sites of glutamate release, supports this hypothesis. However, testing it has been difficult because mechanisms controlling subsynaptic organization remain unknown. Reasoning that transcellular interactions could position AMPA receptors (AMPARs), we targeted a key transsynaptic adhesion molecule implicated in controlling AMPAR number, LRRTM2, using engineered, rapid proteolysis. Severing the LRRTM2 extracellular domain led quickly to nanoscale declustering of AMPARs away from release sites, not prompting their escape from synapses until much later. This rapid remodeling of AMPAR position produced significant deficits in evoked, but not spontaneous, postsynaptic receptor activation. These results dissociate receptor numbers from their nanopositioning in determination of synaptic function and support the novel concept that adhesion molecules acutely position receptors to dynamically control synaptic strength.
Copyright © 2021 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works. Distributed under a Creative Commons Attribution NonCommercial License 4.0 (CC BY-NC).
Figures




References
Grants and funding
LinkOut - more resources
Full Text Sources
