

Vocal learning as a preadaptation for the evolution of human beat perception and synchronization
- PMID: 34420384
- PMCID: PMC8380969
- DOI: 10.1098/rstb.2020.0326
Vocal learning as a preadaptation for the evolution of human beat perception and synchronization
Abstract
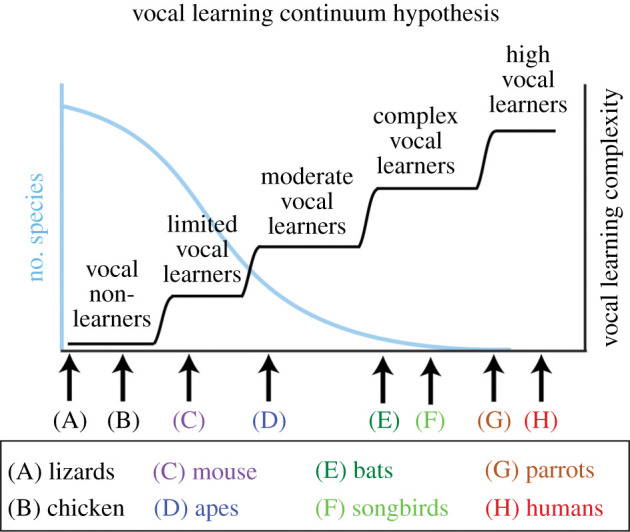
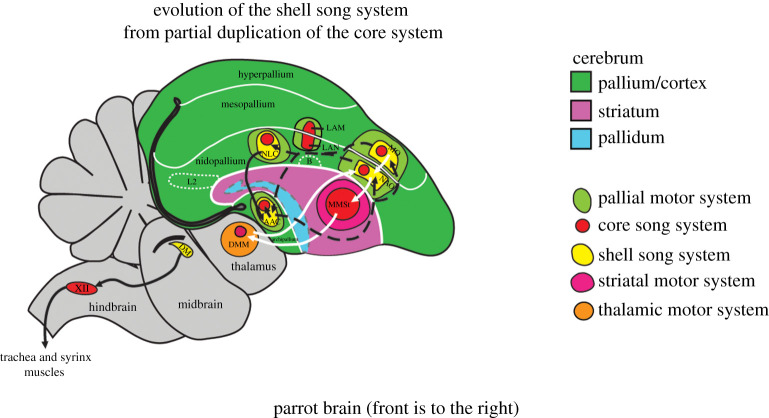
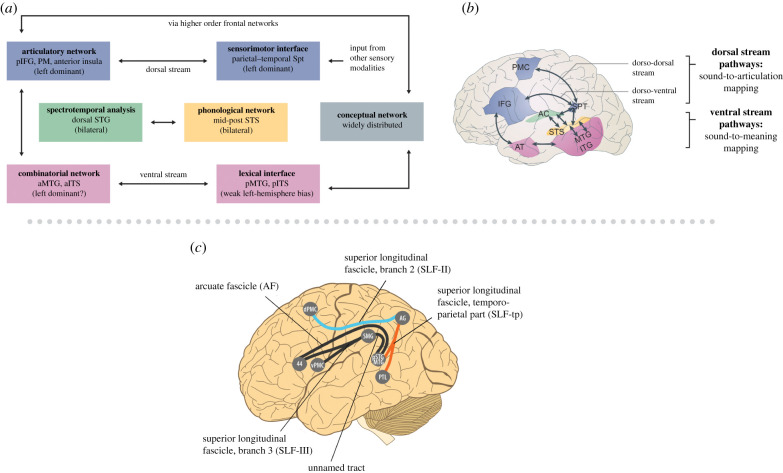
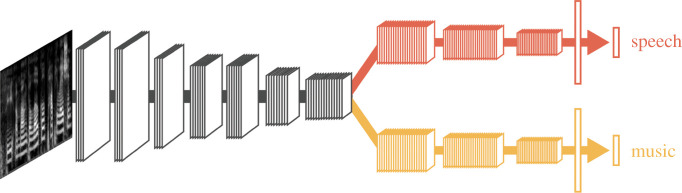
The human capacity to synchronize movements to an auditory beat is central to musical behaviour and to debates over the evolution of human musicality. Have humans evolved any neural specializations for music processing, or does music rely entirely on brain circuits that evolved for other reasons? The vocal learning and rhythmic synchronization hypothesis proposes that our ability to move in time with an auditory beat in a precise, predictive and tempo-flexible manner originated in the neural circuitry for complex vocal learning. In the 15 years, since the hypothesis was proposed a variety of studies have supported it. However, one study has provided a significant challenge to the hypothesis. Furthermore, it is increasingly clear that vocal learning is not a binary trait animals have or lack, but varies more continuously across species. In the light of these developments and of recent progress in the neurobiology of beat processing and of vocal learning, the current paper revises the vocal learning hypothesis. It argues that an advanced form of vocal learning acts as a preadaptation for sporadic beat perception and synchronization (BPS), providing intrinsic rewards for predicting the temporal structure of complex acoustic sequences. It further proposes that in humans, mechanisms of gene-culture coevolution transformed this preadaptation into a genuine neural adaptation for sustained BPS. The larger significance of this proposal is that it outlines a hypothesis of cognitive gene-culture coevolution which makes testable predictions for neuroscience, cross-species studies and genetics. This article is part of the theme issue 'Synchrony and rhythm interaction: from the brain to behavioural ecology'.
Keywords: beat; evolution; gene-culture coevolution; rhythm; synchrony; vocal learning.
Figures
References
-
- Patel, A D. 2008. Music, language, and the brain. New York, NY: Oxford University Press.
-
- Nettl B. 2015. The study of ethnomusicology: thirty-three discussions, 3rd edn. Urbana, IL: University of Illinois Press.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources