

doi: 10.1021/jacs.1c05952.
Epub 2021 Aug 23.
Radicals in Biology: Your Life Is in Their Hands
Affiliations
- PMID: 34423635
- PMCID: PMC8735831
- DOI: 10.1021/jacs.1c05952
Item in Clipboard
Radicals in Biology: Your Life Is in Their Hands
J Am Chem Soc.
.
Abstract
Radicals in biology, once thought to all be bad actors, are now known to play a central role in many enzymatic reactions. Of the known radical-based enzymes, ribonucleotide reductases (RNRs) are pre-eminent as they are essential in the biology of all organisms by providing the building blocks and controlling the fidelity of DNA replication and repair. Intense examination of RNRs has led to the development of new tools and a guiding framework for the study of radicals in biology, pointing the way to future frontiers in radical enzymology.
Figures
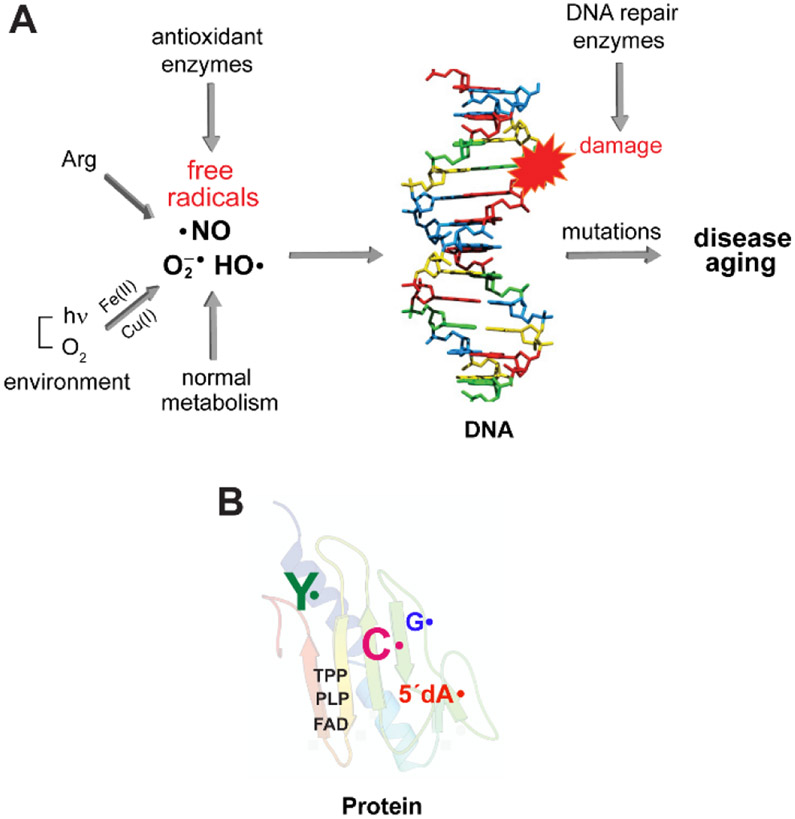
(A) Reactive oxygen radicals such as O2•−/HO• are often generated as the inadvertent consequence of our external or intracellular environments. While O2 is stable, in the presence of Fe(II) and Cu(I), ROS are produced. NO• is produced intracellularly by NO synthase. Its reaction with O2 or O2•− generate RNS. Both ROS and RNS if left to their own demise, can modify DNA, proteins, and lipids and contribute to the aging process and diseased states. (B) In contrast, Nature harnesses the reactivity of radicals to mediate challenging, essential reactions in biology with exquisite specificity. Enzymes involved in primary and secondary metabolism often require cofactors such as flavins (FAD), thiamin pyrophosphate (TPP), pyridoxal phosphate (PLP) that can be involved in controlled radical-based reactions. Stable and transient amino acid radicals of tyrosine (Y•), glycine (G•), cysteine (C•) and 5′-deoxyadenosine (5′-dA•) play essential roles in ribonucleotide reductases (RNRs). These enzymes make the deoxynucleotides essential for DNA replication and are central in the fidelity of the damaged DNA repair. The RNRs and their associated amino acid radicals are the focus of this Perspective.
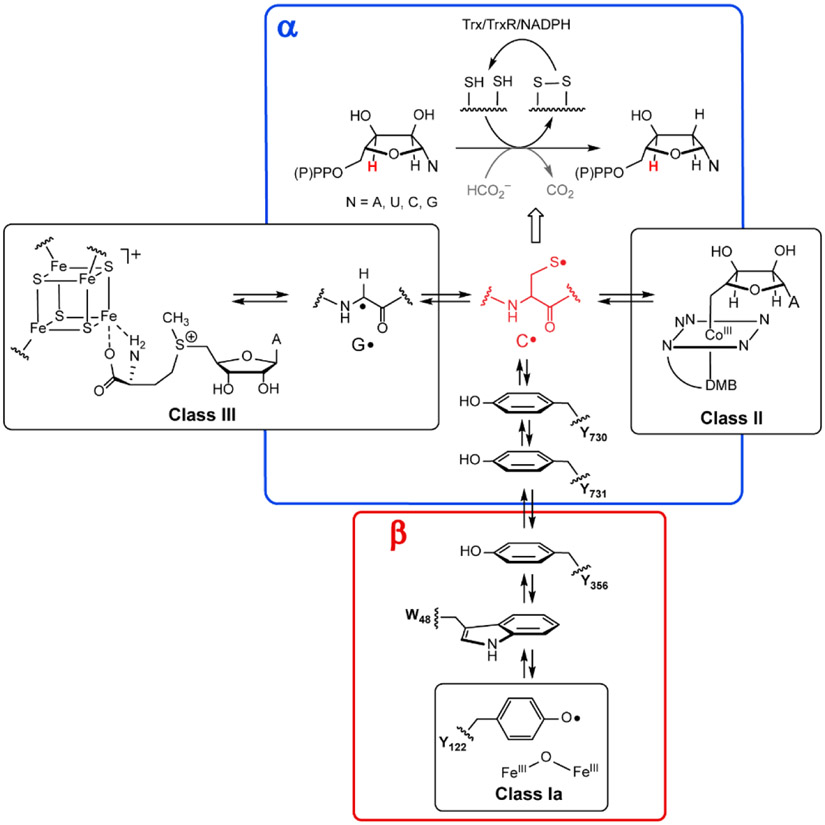
RNRs catalyze de novo deoxynucleoside (di or triphosphates) biosynthesis in all organisms initiated by a cysteine thiyl radical, red C•, in the protein α. It is generated by distinct metallocofactors that define the three classes of RNR (I, II and III). In class I, this process requires a second subunit β (which is part of a homodimer β2) with the metallocofactor initiating C• formation over 32 Å. This process involves a reversible RT pathway with multiple PCET steps involving 4Ys (122, 356, 731, 730), a C (439) and possibly a tryptophan (W48). Residue numbers are for E. coli Ia RNR. The wavy lines indicate that the amino acid radicals are part of the protein. The class II RNRs use adenosylcoblamin (AdoCbl, with a DMB axial ligand(dimethylbenzimidazole). The class III RNRs also require a second protein, the activating enzyme, that uses S-adenosylmethionine and an [4Fe4S] + cluster to generate its glycyl radical (G•). The G• then generates the C• in α. In general, the reducing equivalents to make the deoxynucleotides are supplied by oxidation of two additional cysteines in α to a disulfide. They are recycled by reducing equivalents supplied by redoxin systems such as thioredoxin (Trx), thioredoxin reductase (TrxR) and NADPH. In a subclass of class III RNRs, formate is oxidized to CO2 and supplies the reducing equivalents (gray).
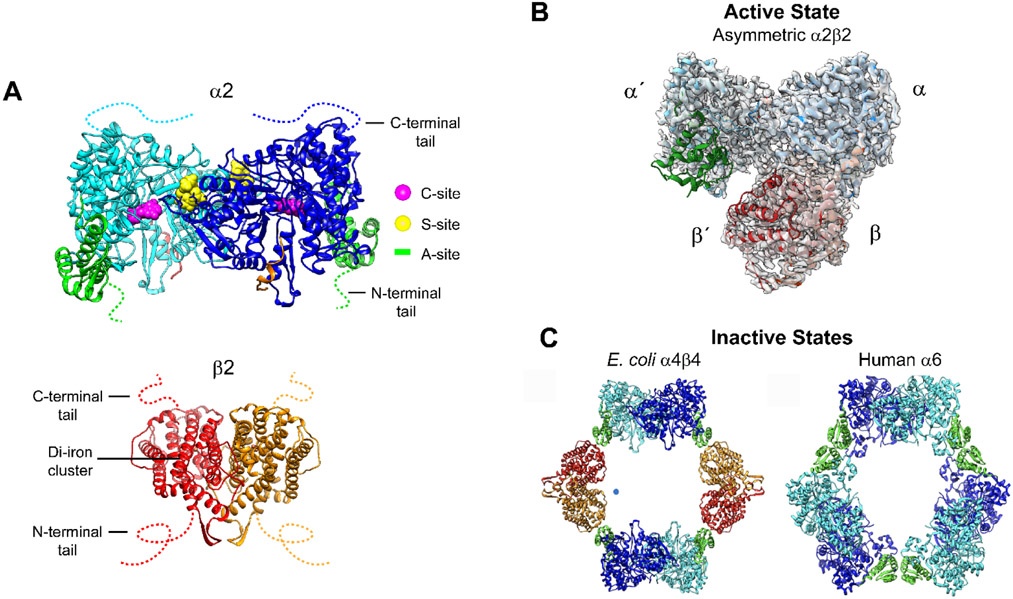
(A) The α (cyan/blue) and β (red/orange) subunits in the E. coli class Ia RNRs are each homodimers. α has three nucleotide binding sites: the catalytic site (C-site), the specificity site (S-site) and the activity site (A site or cone domain, green) in α2, and the radical initiation metallocofactor in β2. The C- and N-termini tails are disordered, indicated by colored dashed lines. (B) The only RNR structure of an active α2β2 complex is an E. coli double mutant that has been trapped in the presence of substrate and effector and solved by cryo-EM. C Structures of dATP inhibited states α4β4 in E. coli and α6 in human Ia RNRs have been solved crystallographically and by cryo-EM.
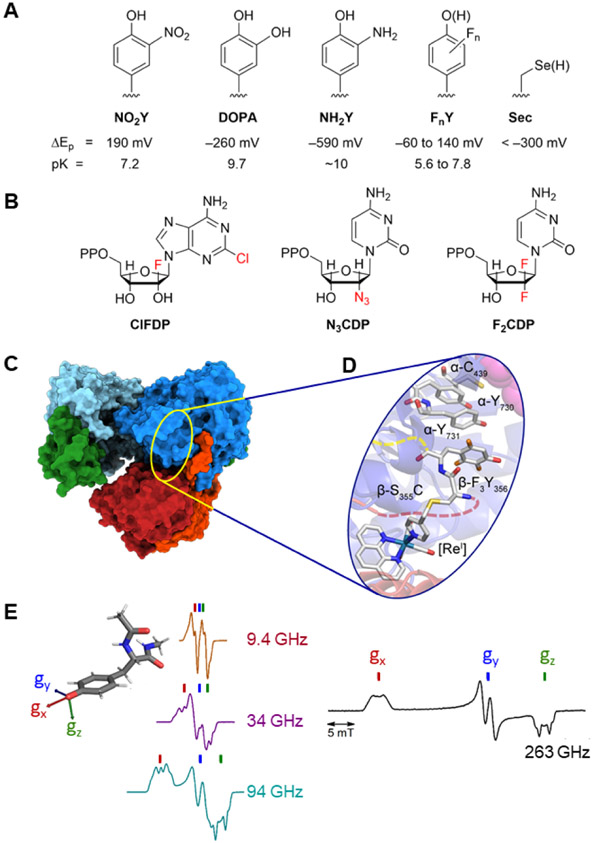
(A) site-specific incorporation of UAA; (B) mechanism-based inhibitors; (C) cyro-EM; (D) radical photogeneration with a covalently attached photooxidant (PO, a [ReI] complex bound to C355 in β; and, (E) high field and multifrequency (9.4, 34, 94, 263 GHz) paramagnetic resonance methods (figure adapted from multifrequency studies of Bennati and coworkers,,).
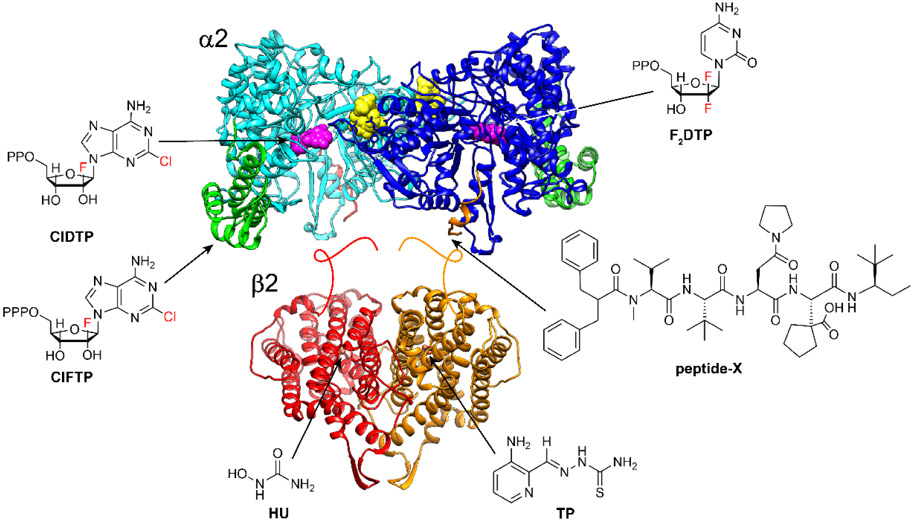
Chlofarabine (ClF) as the diphosphate (ClFDP) and ClFTP reversibly bind to the C-site and A-site of α respectively resulting in formation of “stable” inhibited α6 states. Gemcitabine (F2C) as F2CDP binds to the C-site of α and requires the presence of β to form an active RNR and become a mechanism-based, irreversible inhibitor. Both ClF and F2C are used clinically. Hydroxyurea (HU) and triapine (TP) target β and interfere with the assembly and/or stability of its diferric-tyrosyl radical cofactor. HU is used clinically and triapine is in ongoing clinical trials. The peptide-X analog targets uniquely the herpes simplex virus (HSV) RNR, by specifically binding to its α, interfering with α/β subunit interactions and thus active HSV RNR formation.
References
-
- Winterbourn CC Reconciling the Chemistry and Biology of Reactive Oxygen Species. Nat. Chem. Biol 2008, 4, 278–286. - PubMed
-
- Frey PA; Reed GH The Ubiquity of Iron. ACS Chem. Biol 2012, 7, 1477–1481. - PubMed
-
- Anbar AD Elements and Evolution. Science 2008, 322, 1481–1484. - PubMed
-
- Burney S; Caulfield JL; Niles JR; Wishnok JS; Tannebaum SR The Chemistry of DNA Damage from Nitric Oxide and Peroxynitrite. Mut. Res. – Found. Mol. Mech. Mut 1999, 424, 23–49. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
