

Association of circulating PLA2G7 levels with cancer cachexia and assessment of darapladib as a therapy
- PMID: 34427055
- PMCID: PMC8517355
- DOI: 10.1002/jcsm.12758
Association of circulating PLA2G7 levels with cancer cachexia and assessment of darapladib as a therapy
Abstract
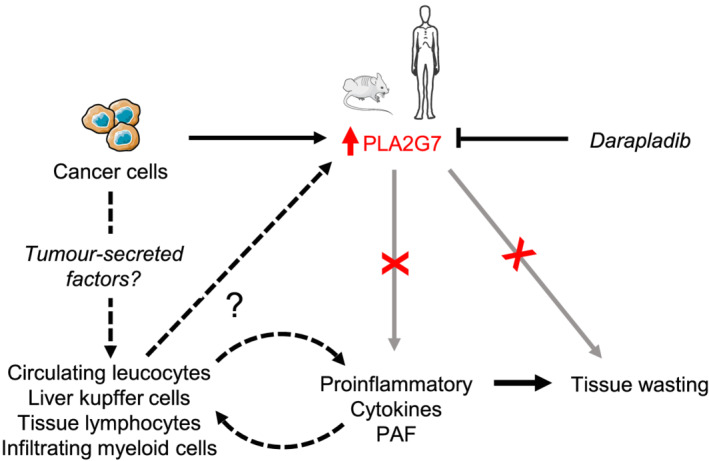
Background: Cancer cachexia (CCx) is a multifactorial wasting disorder characterized by involuntary loss of body weight that affects many cancer patients and implies a poor prognosis, reducing both tolerance to and efficiency of anticancer therapies. Actual challenges in management of CCx remain in the identification of tumour-derived and host-derived mediators involved in systemic inflammation and tissue wasting and in the discovery of biomarkers that would allow for an earlier and personalized care of cancer patients. The aim of this study was to identify new markers of CCx across different species and tumour entities.
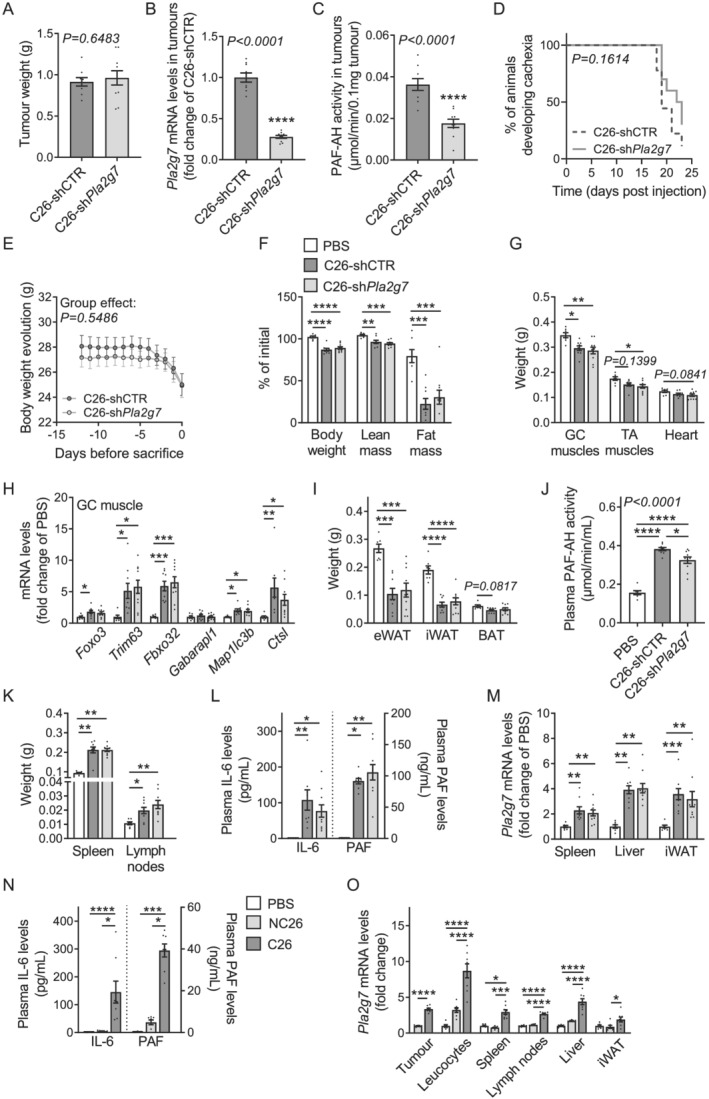
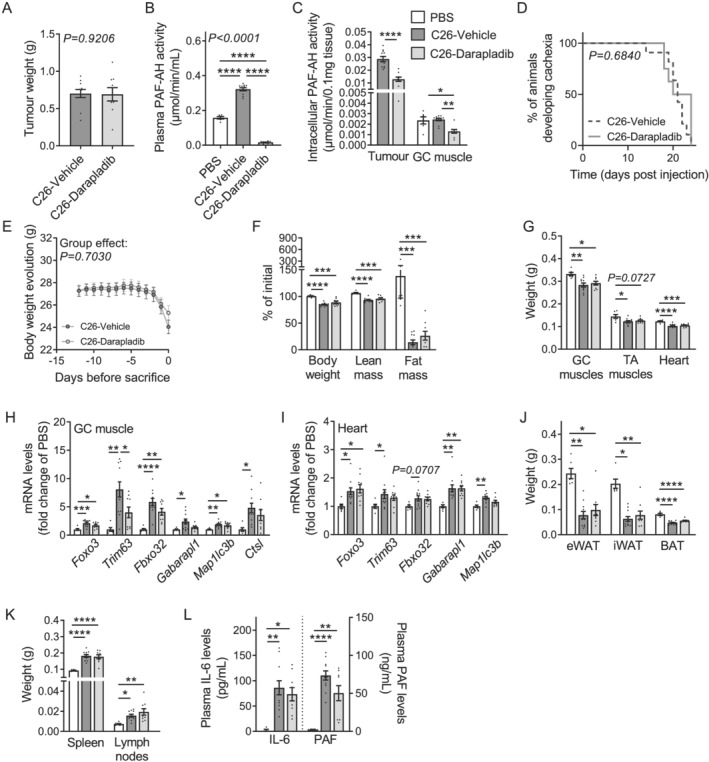
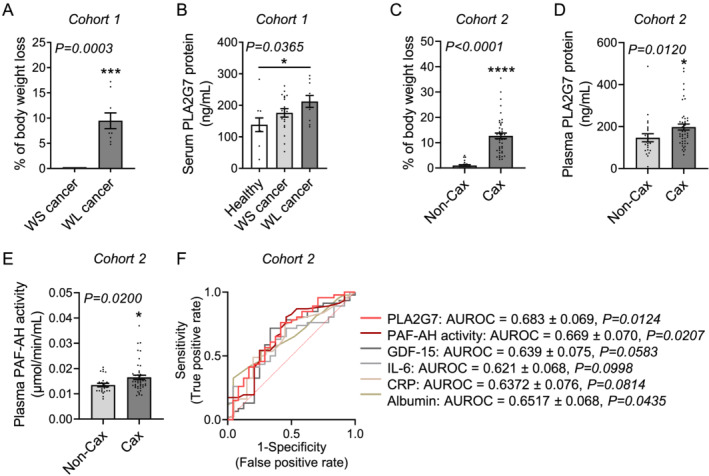
Methods: Quantitative secretome analysis was performed to identify specific factors characteristic of cachexia-inducing cancer cell lines. To establish the subsequently identified phospholipase PLA2G7 as a marker of CCx, plasma PLA2G7 activity and/or protein levels were measured in well-established mouse models of CCx and in different cohorts of weight-stable and weight-losing cancer patients with different tumour entities. Genetic PLA2G7 knock-down in tumours and pharmacological treatment using the well-studied PLA2G7 inhibitor darapladib were performed to assess its implication in the pathogenesis of CCx in C26 tumour-bearing mice.
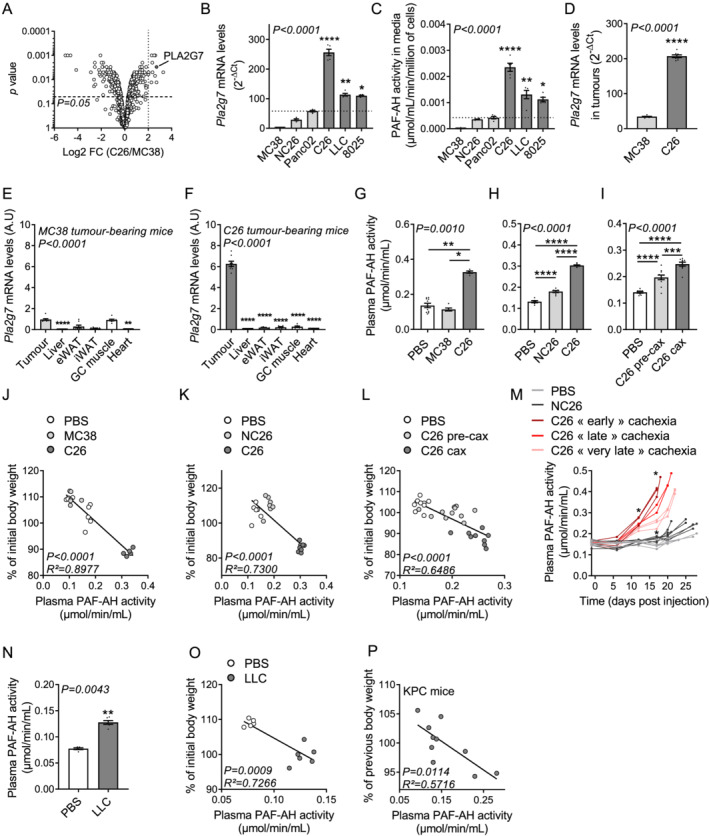
Results: High expression and secretion of PLA2G7 were hallmarks of cachexia-inducing cancer cell lines. Circulating PLA2G7 activity was increased in different mouse models of CCx with various tumour entities and was associated with the severity of body wasting. Circulating PLA2G7 levels gradually rose during cachexia development. Genetic PLA2G7 knock-down in C26 tumours only partially reduced plasma PLA2G7 levels, suggesting that the host is also an important contributor. Chronic treatment with darapladib was not sufficient to counteract inflammation and tissue wasting despite a strong inhibition of the circulating PLA2G7 activity. Importantly, PLA2G7 levels were also increased in colorectal and pancreatic cancer patients with CCx.
Conclusions: Overall, our data show that despite no immediate pathogenic role, at least when targeted as a single entity, PLA2G7 is a consistent marker of CCx in both mice and humans. The early increase in circulating PLA2G7 levels in pre-cachectic mice supports future prospective studies to assess its potential as biomarker for cancer patients.
Keywords: Biomarker; Cancer cachexia; Cancer patients; Darapladib; Mouse models; PLA2G7.
© 2021 The Authors. Journal of Cachexia, Sarcopenia and Muscle published by John Wiley & Sons Ltd on behalf of Society on Sarcopenia, Cachexia and Wasting Disorders.
Conflict of interest statement
None declared.
Figures
References
-
- Baracos VE, Martin L, Korc M, Guttridge DC, Fearon KC. Cancer‐associated cachexia. Nat Rev Dis Primers 2018;4:17105. - PubMed
-
- Fearon K, Strasser F, Anker SD, Bosaeus I, Bruera E, Fainsinger RL, et al. Definition and classification of cancer cachexia: an international consensus. Lancet Oncol 2011;12:489–495. - PubMed
-
- Argiles JM, Stemmler B, López‐Soriano FJ, Busquets S. Inter‐tissue communication in cancer cachexia. Nat Rev Endocrinol 2018;15:9–20. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
