

Metabolic plasticity drives development during mammalian embryogenesis
- PMID: 34428399
- PMCID: PMC8584261
- DOI: 10.1016/j.devcel.2021.07.020
Metabolic plasticity drives development during mammalian embryogenesis
Abstract
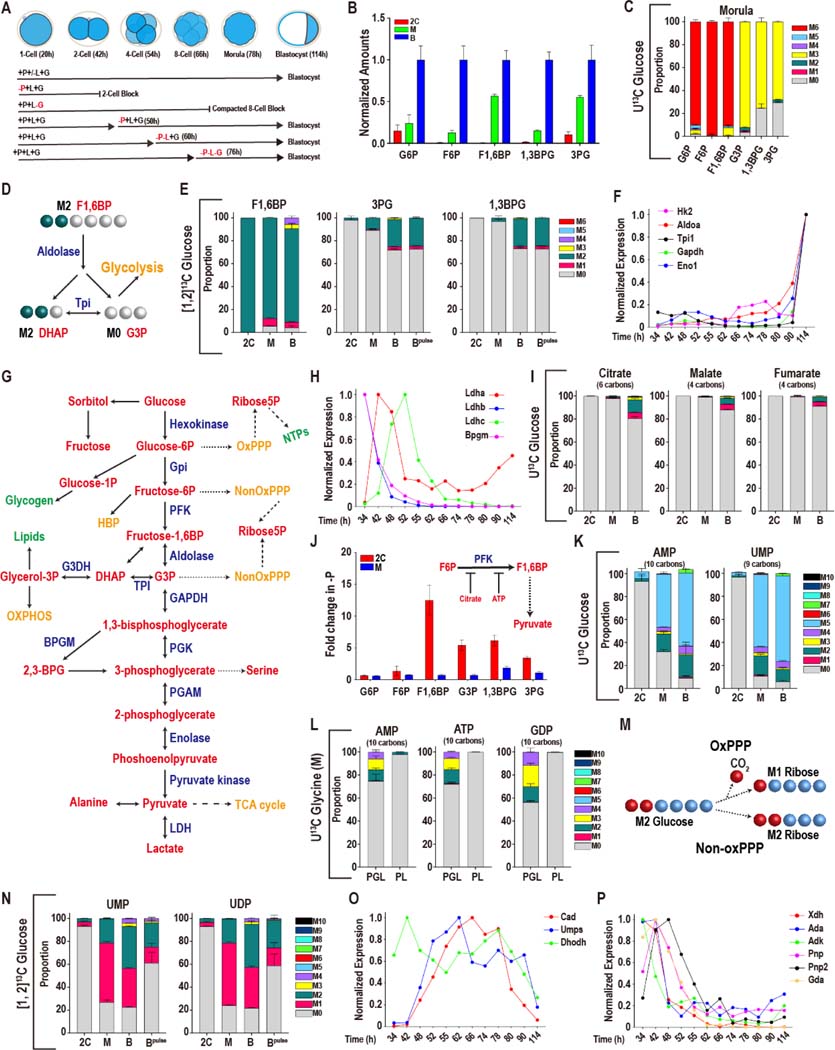
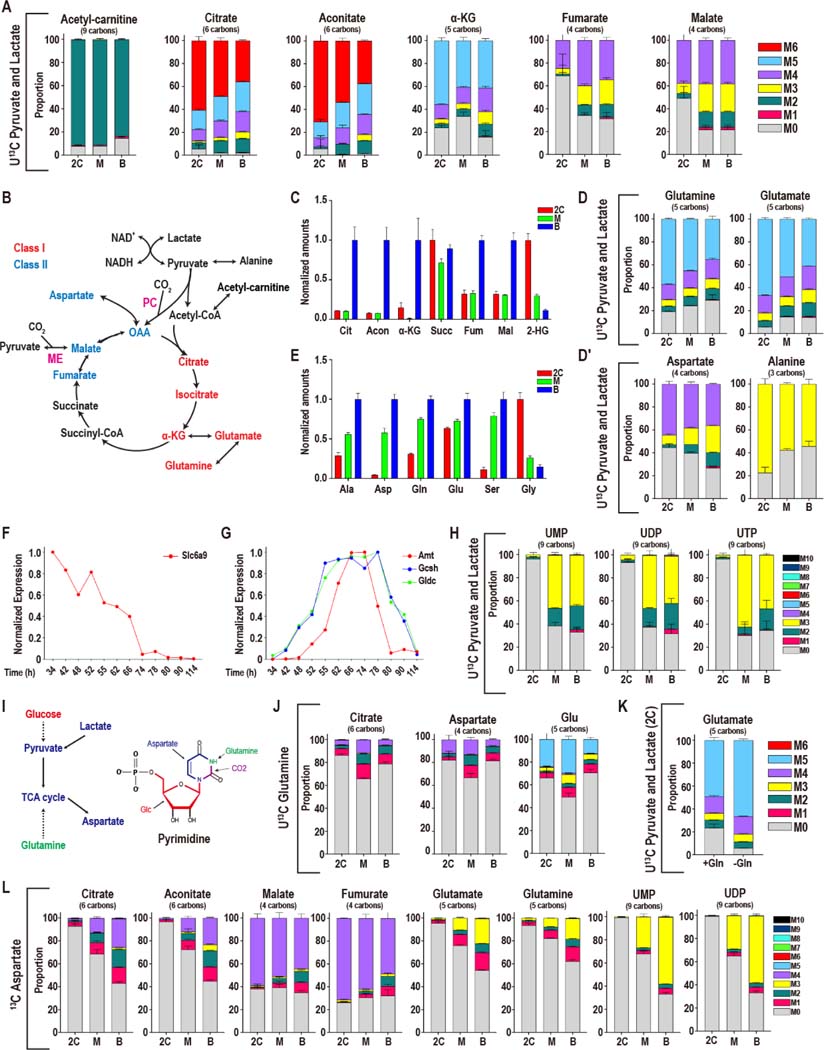
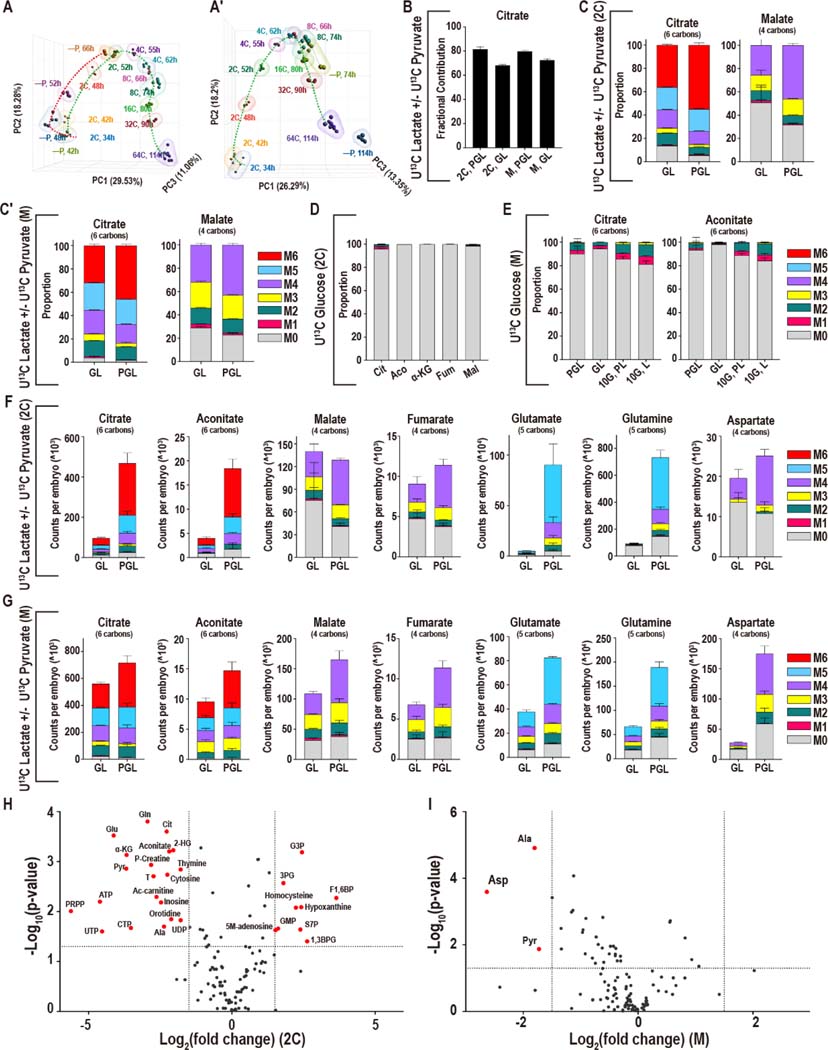
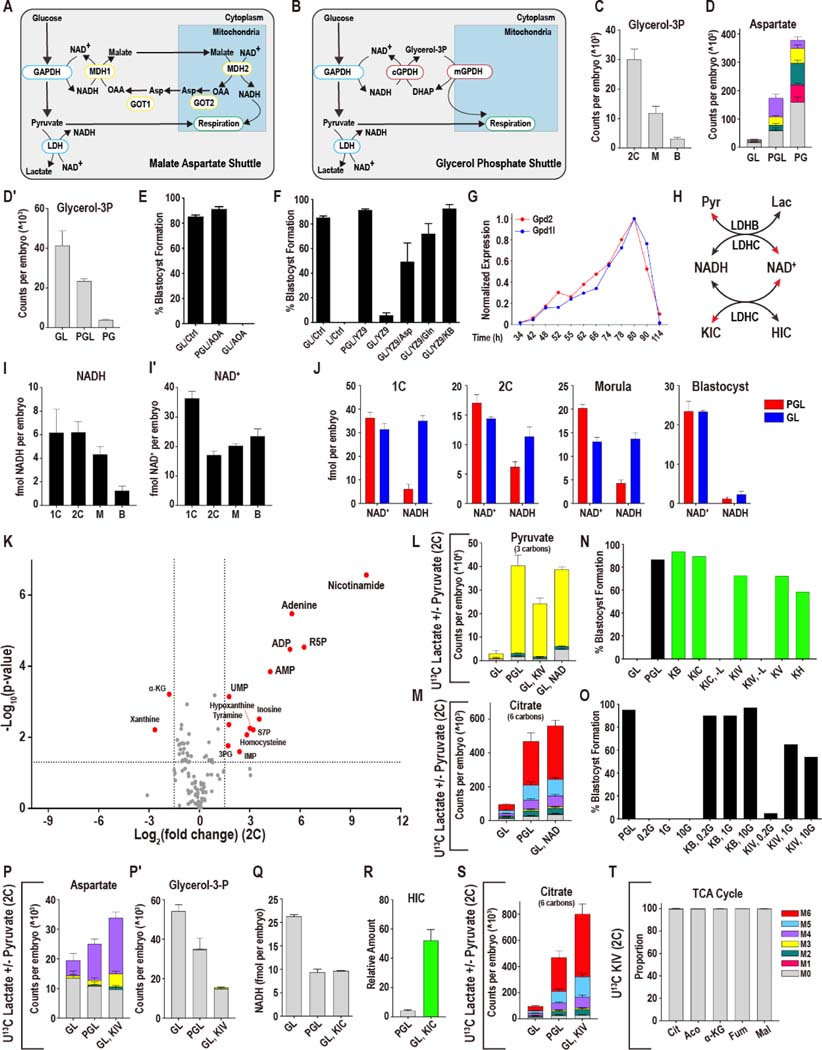
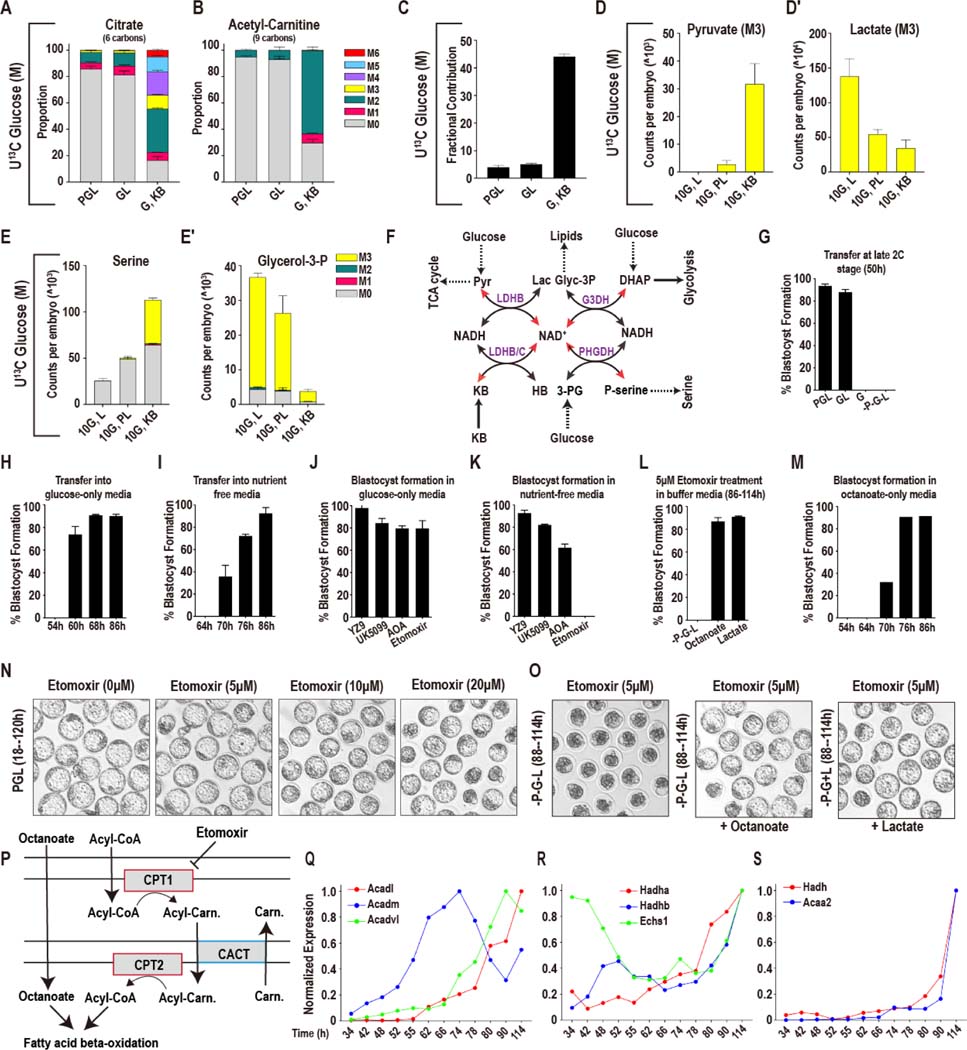
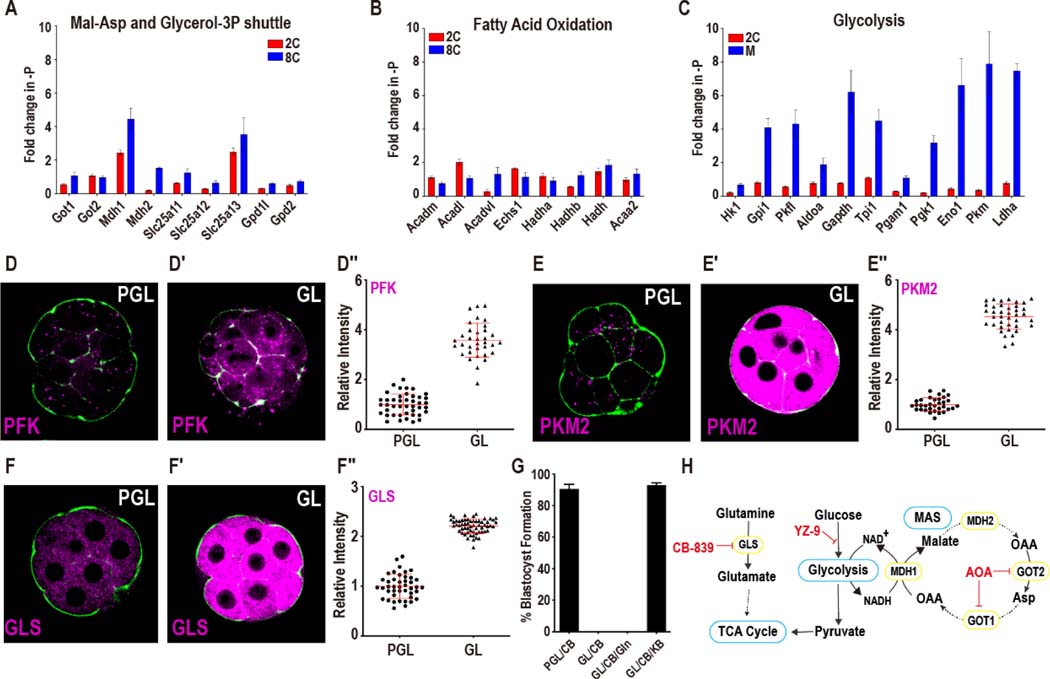
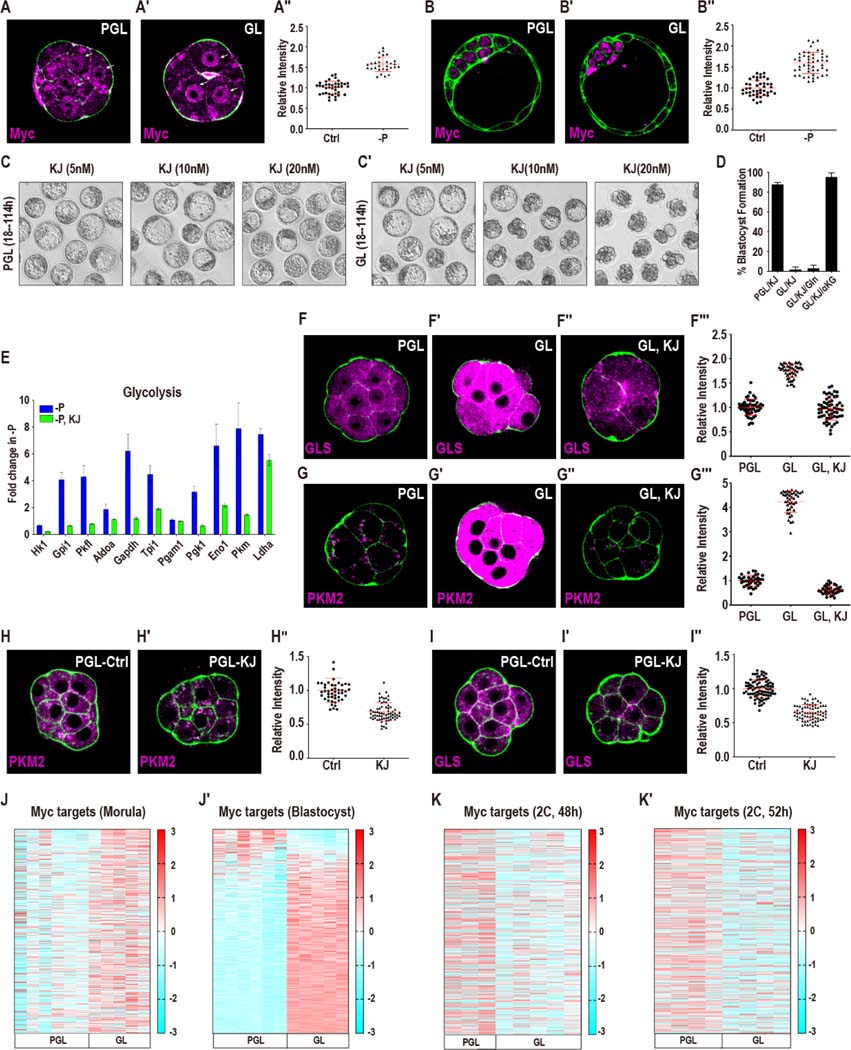
Mammalian preimplantation embryos follow a stereotypic pattern of development from zygotes to blastocysts. Here, we use labeled nutrient isotopologue analysis of small numbers of embryos to track downstream metabolites. Combined with transcriptomic analysis, we assess the capacity of the embryo to reprogram its metabolism through development. Early embryonic metabolism is rigid in its nutrient requirements, sensitive to reductive stress and has a marked disequilibrium between two halves of the TCA cycle. Later, loss of maternal LDHB and transcription of zygotic products favors increased activity of bioenergetic shuttles, fatty-acid oxidation and equilibration of the TCA cycle. As metabolic plasticity peaks, blastocysts can develop without external nutrients. Normal developmental metabolism of the early embryo is distinct from cancer metabolism. However, similarities emerge upon reductive stress. Increased metabolic plasticity with maturation is due to changes in redox control mechanisms and to transcriptional reprogramming of later-stage embryos during homeostasis or upon adaptation to environmental changes.
Keywords: MYC; NAD+/NADH; developmental metabolism; embryo; metabolic plasticity; metabolic reprogramming; preimplantation; redox; reductive stress; zygotic genome activation.
Copyright © 2021 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
References
-
- Alexiou M, and Leese HJ (1992). Purine utilisation, de novo synthesis and degradation in mouse preimplantation embryos. Development 114, 185–192. - PubMed
-
- Alexiou M, and Leese HJ (1994). Enzymes of purine salvage and catabolism in the mouse preimplantation embryo measured by high performance liquid chromatography. J Reprod Fertil 101, 151–158. - PubMed
-
- Altman BJ, Stine ZE, and Dang CV (2016). From Krebs to clinic: glutamine metabolism to cancer therapy. Nat Rev Cancer 16, 749. - PubMed
-
- Aragon JJ, Tornheim K, Goodman MN, and Lowenstein JM (1981). Replenishment of citric acid cycle intermediates by the purine nucleotide cycle in rat skeletal muscle. Curr Top Cell Regul 18, 131–149. - PubMed
-
- Baltz JM (2001). Osmoregulation and cell volume regulation in the preimplantation embryo. Curr Top Dev Biol 52, 55–106. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
