

Systematic Profiling of DNMT3A Variants Reveals Protein Instability Mediated by the DCAF8 E3 Ubiquitin Ligase Adaptor
- PMID: 34429321
- PMCID: PMC8758508
- DOI: 10.1158/2159-8290.CD-21-0560
Systematic Profiling of DNMT3A Variants Reveals Protein Instability Mediated by the DCAF8 E3 Ubiquitin Ligase Adaptor
Abstract
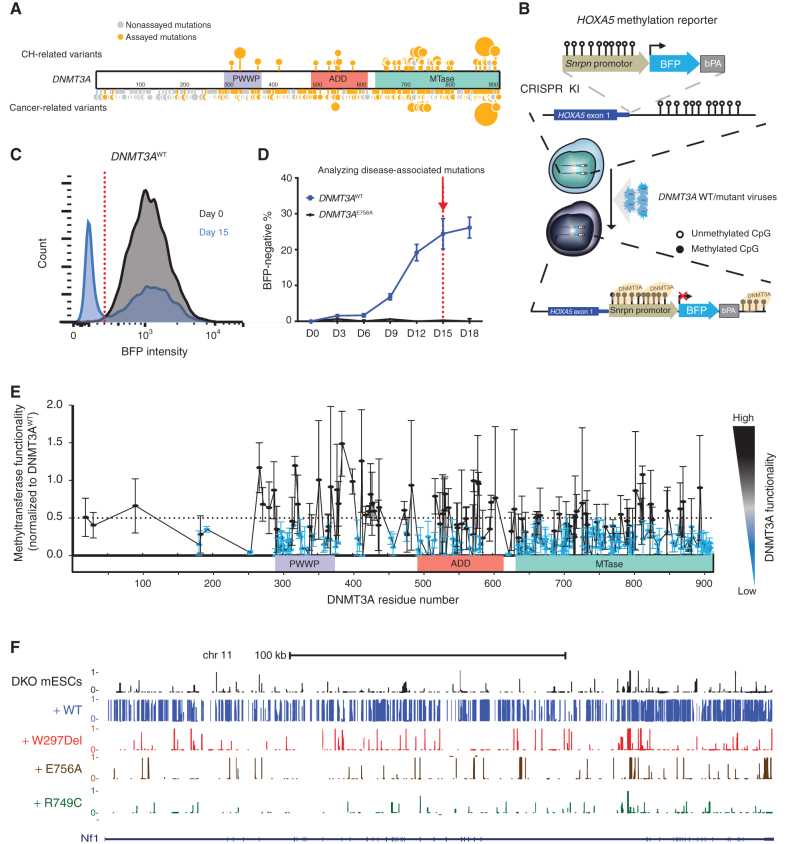
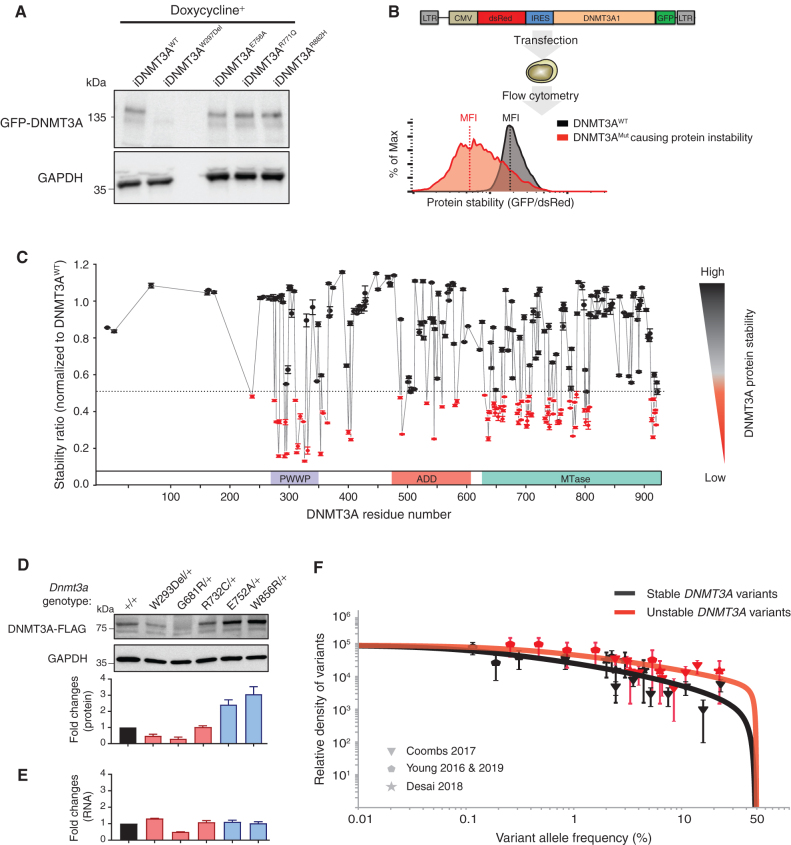
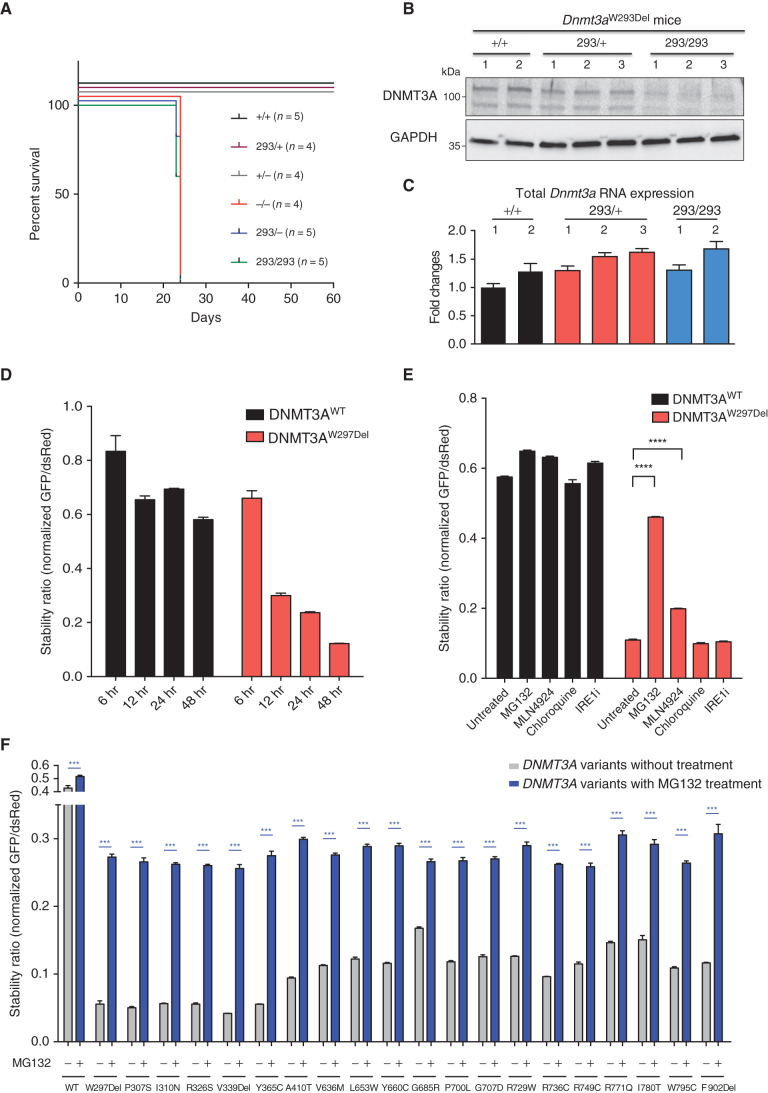
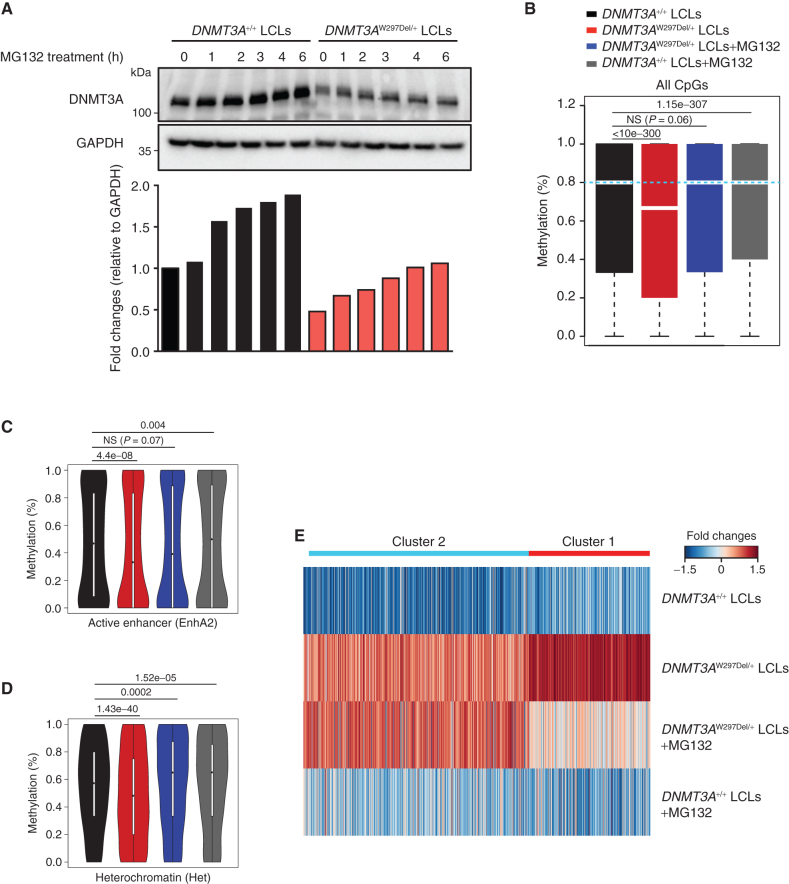
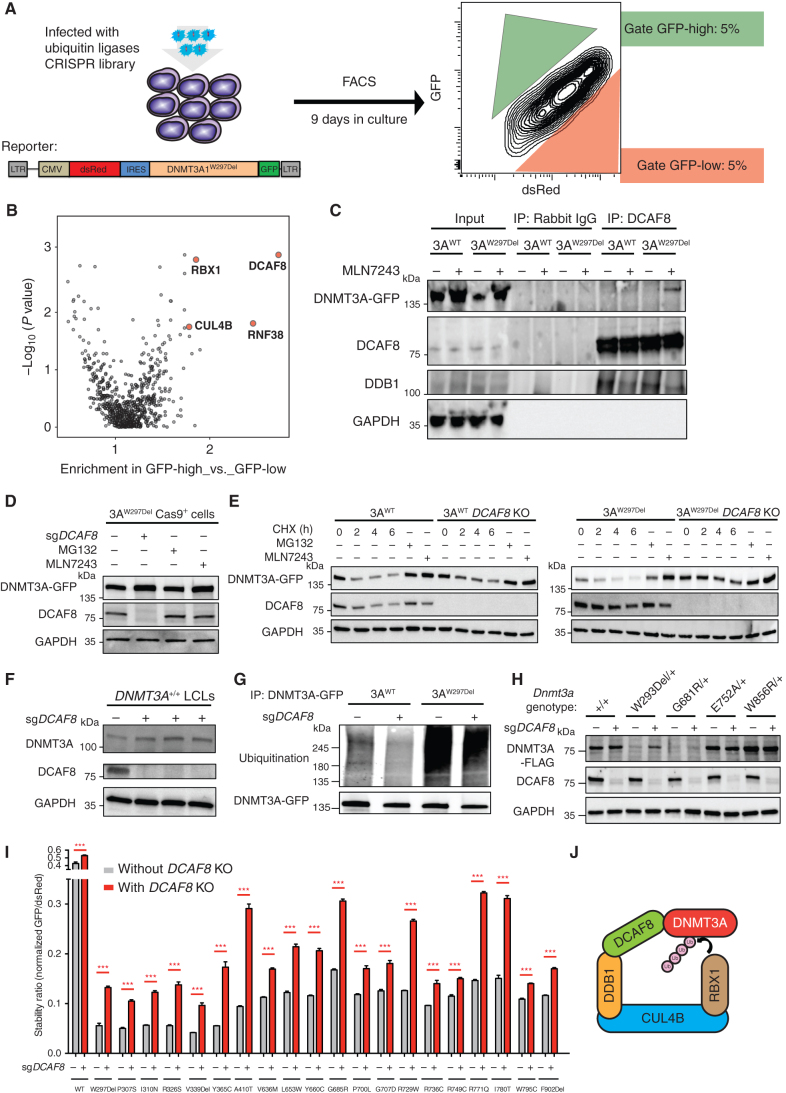
Clonal hematopoiesis is a prevalent age-related condition associated with a greatly increased risk of hematologic disease; mutations in DNA methyltransferase 3A (DNMT3A) are the most common driver of this state. DNMT3A variants occur across the gene with some particularly associated with malignancy, but the functional relevance and mechanisms of pathogenesis of the majority of mutations are unknown. Here, we systematically investigated the methyltransferase activity and protein stability of 253 disease-associated DNMT3A mutations, and found that 74% were loss-of-function mutations. Half of these variants exhibited reduced protein stability and, as a class, correlated with greater clonal expansion and acute myeloid leukemia development. We investigated the mechanisms underlying the instability using a CRISPR screen and uncovered regulated destruction of DNMT3A mediated by the DCAF8 E3 ubiquitin ligase adaptor. We establish a new paradigm to classify novel variants that has prognostic and potential therapeutic significance for patients with hematologic disease. SIGNIFICANCE: DNMT3A has emerged as the most important epigenetic regulator and tumor suppressor in the hematopoietic system. Our study represents a systematic and high-throughput method to characterize the molecular impact of DNMT3A missense mutations and the discovery of a regulated destruction mechanism of DNMT3A offering new prognostic and future therapeutic avenues.See related commentary by Ma and Will, p. 23.This article is highlighted in the In This Issue feature, p. 1.
©2021 The Authors; Published by the American Association for Cancer Research.
Figures
Comment in
-
To Degrade or Not to Degrade DNMT3A.Cancer Discov. 2022 Jan;12(1):23-25. doi: 10.1158/2159-8290.CD-21-1430. Cancer Discov. 2022. PMID: 35022208
References
Publication types
MeSH terms
Substances
Grants and funding
- F99 CA222736/CA/NCI NIH HHS/United States
- R56 DK092883/DK/NIDDK NIH HHS/United States
- R01 GM112003/GM/NIGMS NIH HHS/United States
- R01 CA228140/CA/NCI NIH HHS/United States
- 29749/CRUK_/Cancer Research UK/United Kingdom
- S10 RR024574/RR/NCRR NIH HHS/United States
- R01 DK092883/DK/NIDDK NIH HHS/United States
- P30 CA125123/CA/NCI NIH HHS/United States
- UM1 HG006348/HG/NHGRI NIH HHS/United States
- R01 HL134780/HL/NHLBI NIH HHS/United States
- K00 CA222736/CA/NCI NIH HHS/United States
- R01 CA183252/CA/NCI NIH HHS/United States
- R01 HG007538/HG/NHGRI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
