

Depletion of TAX1BP1 Amplifies Innate Immune Responses during Respiratory Syncytial Virus Infection
- PMID: 34431698
- PMCID: PMC8549506
- DOI: 10.1128/JVI.00912-21
Depletion of TAX1BP1 Amplifies Innate Immune Responses during Respiratory Syncytial Virus Infection
Abstract
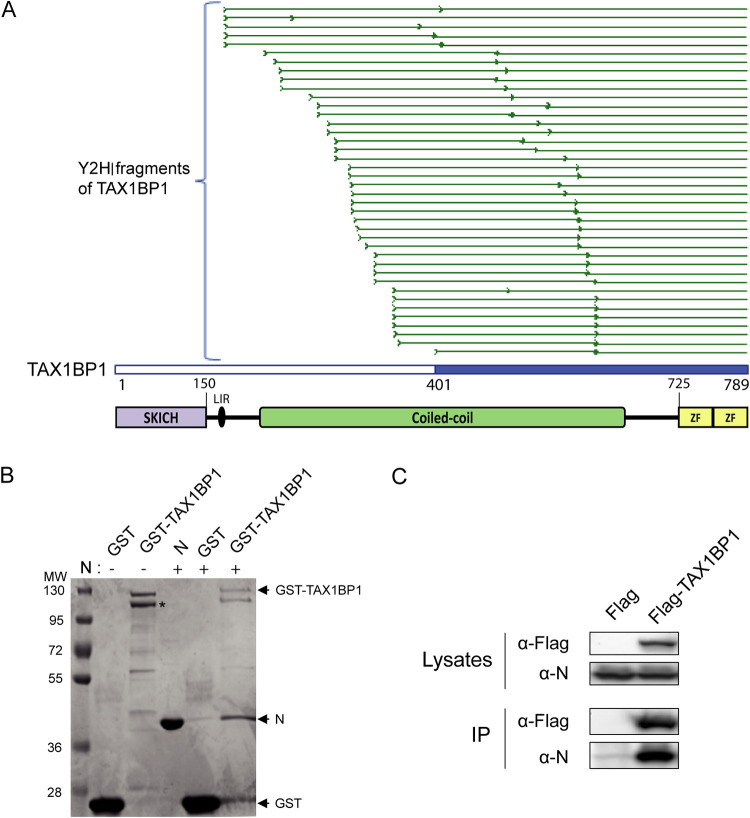
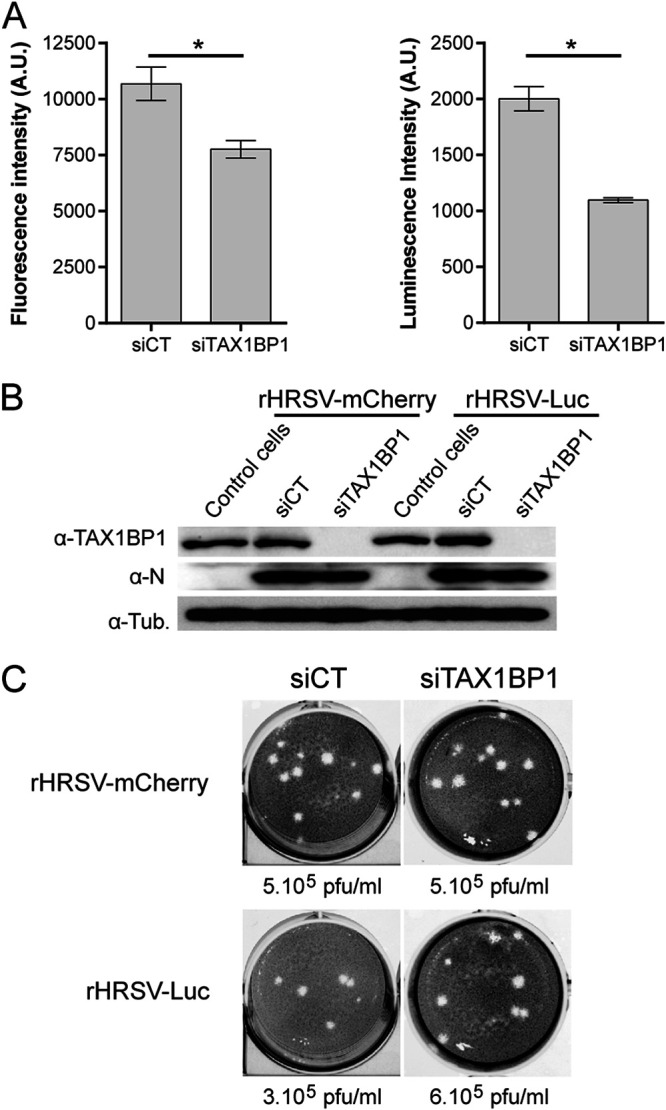
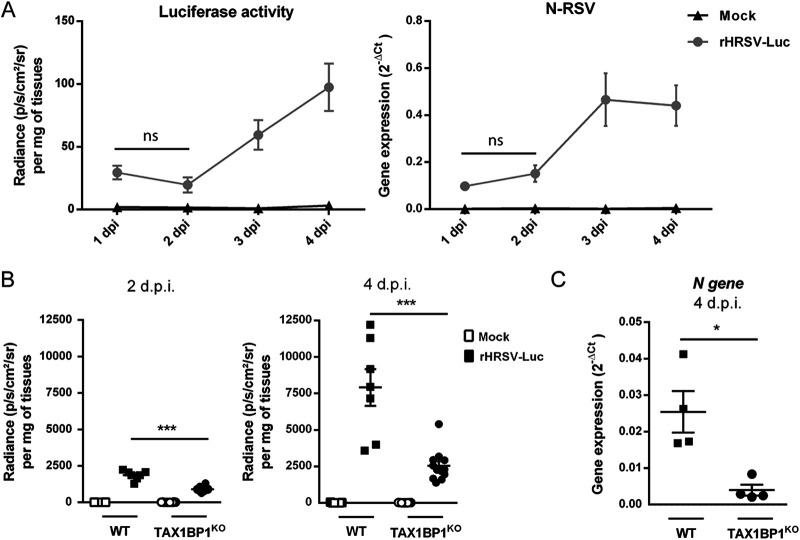
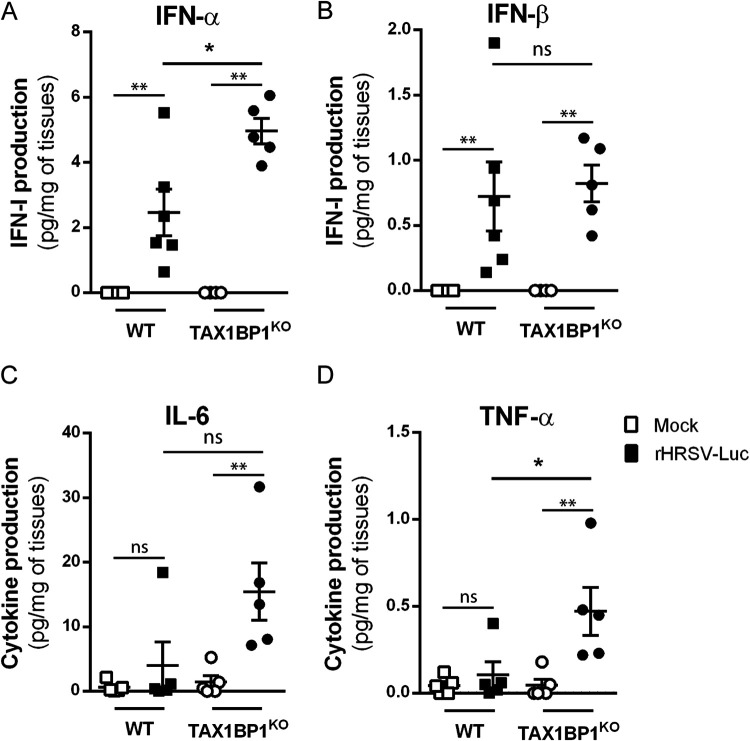
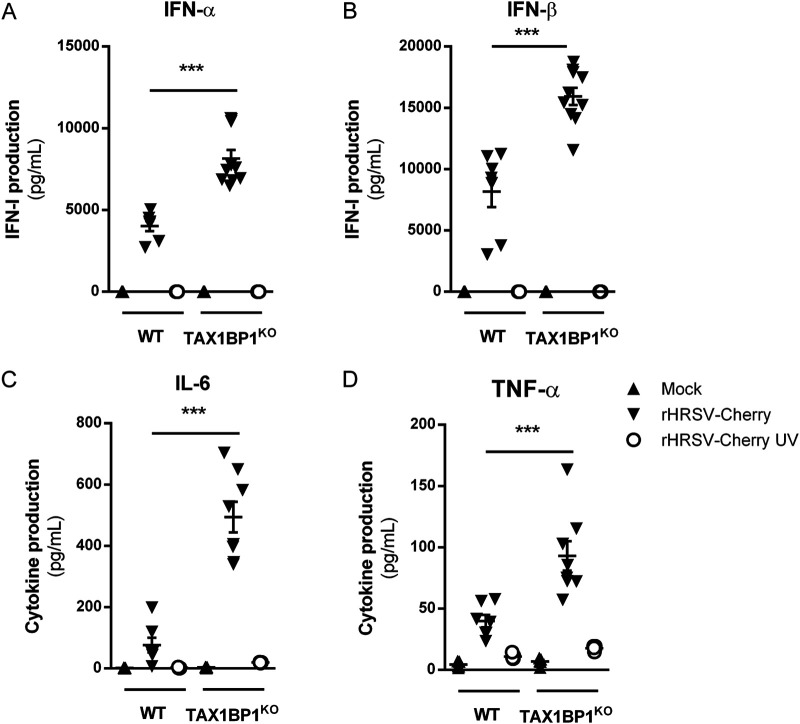
Respiratory syncytial virus (RSV) is the main cause of acute respiratory infections in young children and also has a major impact on the elderly and immunocompromised people. In the absence of a vaccine or efficient treatment, a better understanding of RSV interactions with the host antiviral response during infection is needed. Previous studies revealed that cytoplasmic inclusion bodies (IBs), where viral replication and transcription occur, could play a major role in the control of innate immunity during infection by recruiting cellular proteins involved in the host antiviral response. We recently showed that the morphogenesis of IBs relies on a liquid-liquid-phase separation mechanism depending on the interaction between viral nucleoprotein (N) and phosphoprotein (P). These scaffold proteins are expected to play a central role in the recruitment of cellular proteins to IBs. Here, we performed a yeast two-hybrid screen using RSV N protein as bait and identified the cellular protein TAX1BP1 as a potential partner of this viral protein. This interaction was validated by pulldown and immunoprecipitation assays. We showed that TAX1BP1 suppression has only a limited impact on RSV infection in cell cultures. However, RSV replication is decreased in TAX1BP1-deficient (TAX1BP1 knockout [TAX1BP1KO]) mice, whereas the production of inflammatory and antiviral cytokines is enhanced. In vitro infection of wild-type or TAX1BP1KO alveolar macrophages confirmed that the innate immune response to RSV infection is enhanced in the absence of TAX1BP1. Altogether, our results suggest that RSV could hijack TAX1BP1 to restrain the host immune response during infection. IMPORTANCE Respiratory syncytial virus (RSV), which is the leading cause of lower respiratory tract illness in infants, remains a medical problem in the absence of a vaccine or efficient treatment. This virus is also recognized as a main pathogen in the elderly and immunocompromised people, and the occurrence of coinfections (with other respiratory viruses and bacteria) amplifies the risks of developing respiratory distress. In this context, a better understanding of the pathogenesis associated with viral respiratory infections, which depends on both viral replication and the host immune response, is needed. The present study reveals that the cellular protein TAX1BP1, which interacts with the RSV nucleoprotein N, participates in the control of the innate immune response during RSV infection, suggesting that the N-TAX1BP1 interaction represents a new target for the development of antivirals.
Keywords: RSV; TAX1BP1; innate immunity; interferons; lung; nucleoprotein; yeast two-hybrid screening.
Figures
References
-
- Shi T, McAllister DA, O’Brien KL, Simoes EAF, Madhi SA, Gessner BD, Polack FP, Balsells E, Acacio S, Aguayo C, Alassani I, Ali A, Antonio M, Awasthi S, Awori JO, Azziz-Baumgartner E, Baggett HC, Baillie VL, Balmaseda A, Barahona A, Basnet S, Bassat Q, Basualdo W, Bigogo G, Bont L, Breiman RF, Brooks WA, Broor S, Bruce N, Bruden D, Buchy P, Campbell S, Carosone-Link P, Chadha M, Chipeta J, Chou M, Clara W, Cohen C, de Cuellar E, Dang D-A, Dash-Yandag B, Deloria-Knoll M, Dherani M, Eap T, Ebruke BE, Echavarria M, de Freitas Lazaro Emediato CC, Fasce RA, Feikin DR, Feng L, et al.. 2017. Global, regional, and national disease burden estimates of acute lower respiratory infections due to respiratory syncytial virus in young children in 2015: a systematic review and modelling study. Lancet 390:946–958. 10.1016/S0140-6736(17)30938-8. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
