

Activation of Cph1 causes ß(1,3)-glucan unmasking in Candida albicans and attenuates virulence in mice in a neutrophil-dependent manner
- PMID: 34432857
- PMCID: PMC8423308
- DOI: 10.1371/journal.ppat.1009839
Activation of Cph1 causes ß(1,3)-glucan unmasking in Candida albicans and attenuates virulence in mice in a neutrophil-dependent manner
Abstract
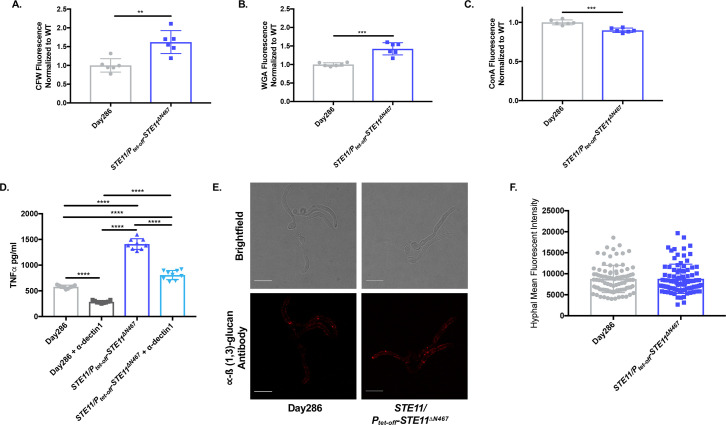
Masking the immunogenic cell wall epitope ß(1,3)-glucan under an outer layer of mannosylated glycoproteins is an important virulence factor deployed by Candida albicans during infection. Consequently, increased ß(1,3)-glucan exposure (unmasking) reveals C. albicans to the host's immune system and attenuates its virulence. We have previously shown that activation of the Cek1 MAPK pathway via expression of a hyperactive allele of an upstream kinase (STE11ΔN467) induced unmasking. It also increased survival of mice in a murine disseminated candidiasis model and attenuated kidney fungal burden by ≥33 fold. In this communication, we utilized cyclophosphamide-induced immunosuppression to test if the clearance of the unmasked STE11ΔN467 mutant was dependent on the host immune system. Suppression of the immune response by cyclophosphamide reduced the attenuation in fungal burden caused by the STE11ΔN467 allele. Moreover, specific depletion of neutrophils via 1A8 antibody treatment also reduced STE11ΔN467-dependent fungal burden attenuation, but to a lesser extent than cyclophosphamide, demonstrating an important role for neutrophils in mediating fungal clearance of unmasked STE11ΔN467 cells. In an effort to understand the mechanism by which Ste11ΔN467 causes unmasking, transcriptomics were used to reveal that several components in the Cek1 MAPK pathway were upregulated, including the transcription factor CPH1 and the cell wall sensor DFI1. In this report we show that a cph1ΔΔ mutation restored ß(1,3)-glucan exposure to wild-type levels in the STE11ΔN467 strain, confirming that Cph1 is the transcription factor mediating Ste11ΔN467-induced unmasking. Furthermore, Cph1 is shown to induce a positive feedback loop that increases Cek1 activation. In addition, full unmasking by STE11ΔN467 is dependent on the upstream cell wall sensor DFI1. However, while deletion of DFI1 significantly reduced Ste11ΔN467-induced unmasking, it did not impact activation of the downstream kinase Cek1. Thus, it appears that once stimulated by Ste11ΔN467, Dfi1 activates a parallel signaling pathway that is involved in Ste11ΔN467-induced unmasking.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures








References
Publication types
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
