

Structured elements drive extensive circular RNA translation
- PMID: 34437836
- PMCID: PMC8567535
- DOI: 10.1016/j.molcel.2021.07.042
Structured elements drive extensive circular RNA translation
Abstract
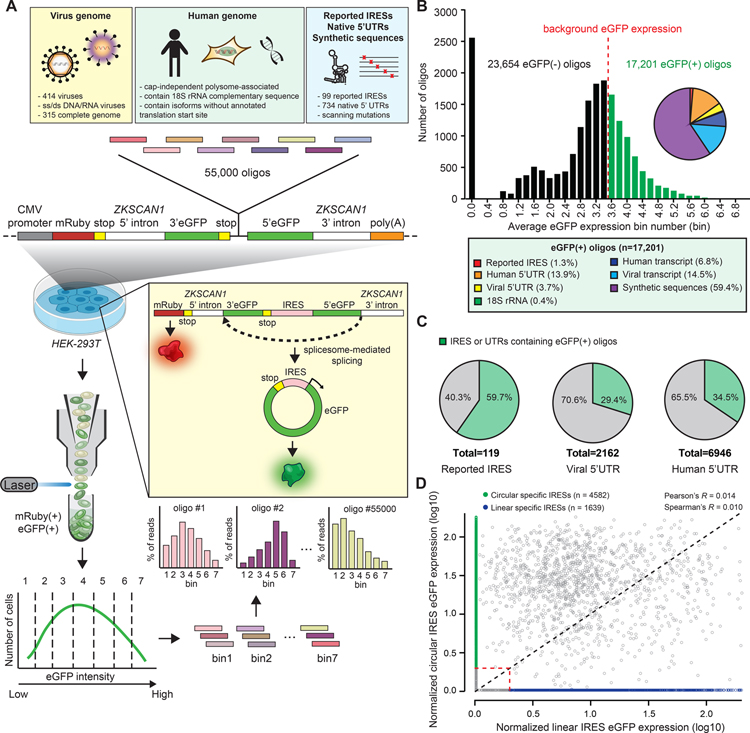
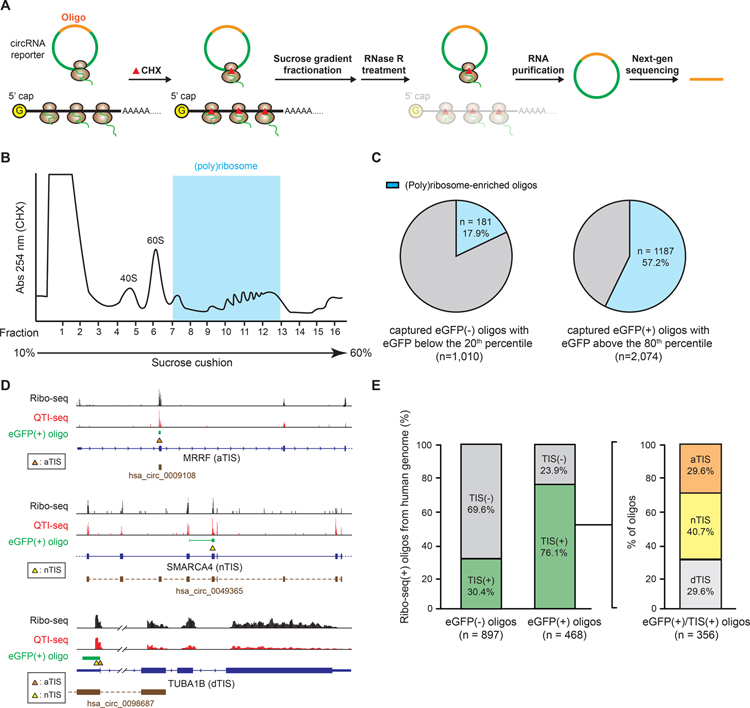
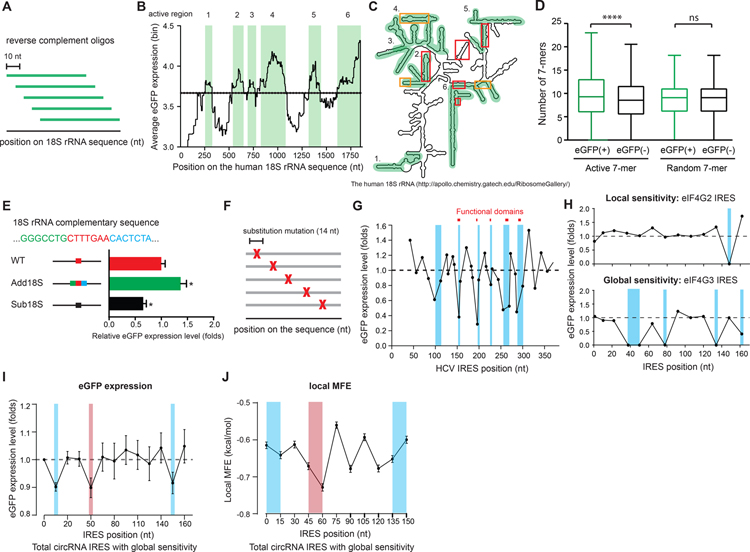
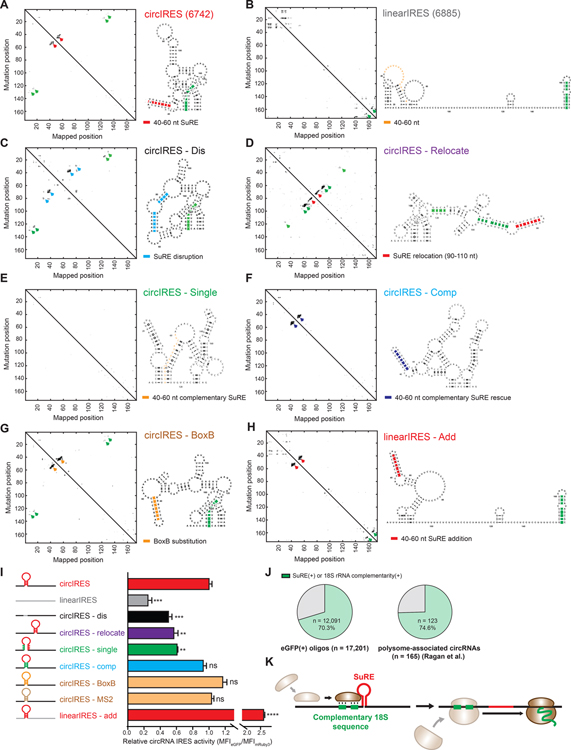
The human genome encodes tens of thousands circular RNAs (circRNAs) with mostly unknown functions. Circular RNAs require internal ribosome entry sites (IRES) if they are to undergo translation without a 5' cap. Here, we develop a high-throughput screen to systematically discover RNA sequences that can direct circRNA translation in human cells. We identify more than 17,000 endogenous and synthetic sequences as candidate circRNA IRES. 18S rRNA complementarity and a structured RNA element positioned on the IRES are important for driving circRNA translation. Ribosome profiling and peptidomic analyses show extensive IRES-ribosome association, hundreds of circRNA-encoded proteins with tissue-specific distribution, and antigen presentation. We find that circFGFR1p, a protein encoded by circFGFR1 that is downregulated in cancer, functions as a negative regulator of FGFR1 oncoprotein to suppress cell growth during stress. Systematic identification of circRNA IRES elements may provide important links among circRNA regulation, biological function, and disease.
Keywords: 18S complementarity; FGFR1; cap-independent translation; circFGFR1p; circRNA-encoded protein; circular RNA; internal ribosome entry site; structured RNA element.
Copyright © 2021 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests Stanford University has filed patent applications on the basis of this work, and H.Y.C. and C.-K.C. are named as co-inventors. H.Y.C. is a co-founder and advisor of Accent Therapeutics, Boundless Bio, Cartography Biosciences, and Circ Bio. H.Y.C. is an advisor of 10X Genomics, Arsenal Biosciences, and Spring Discovery. H.Y.C. is a member of the Molecular Cell advisory board.
Figures
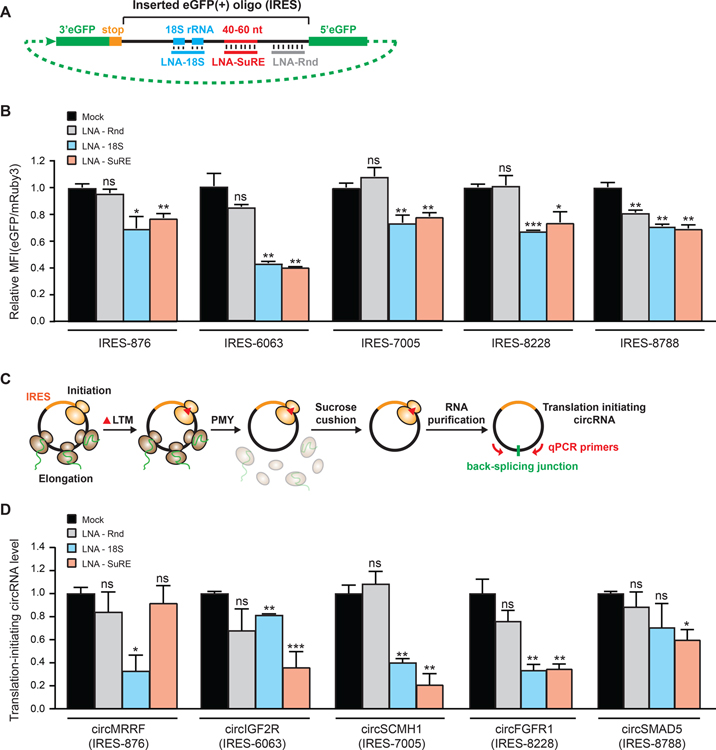
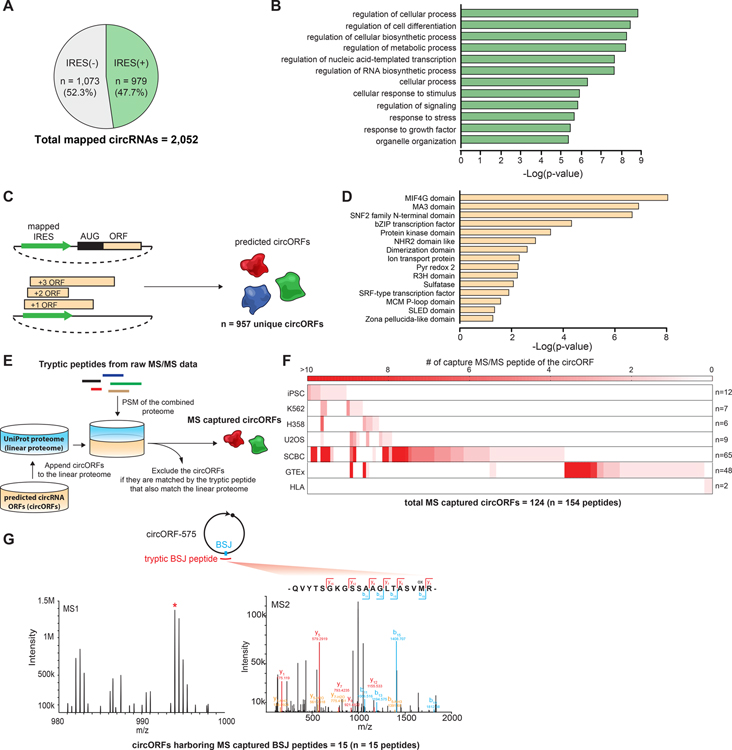
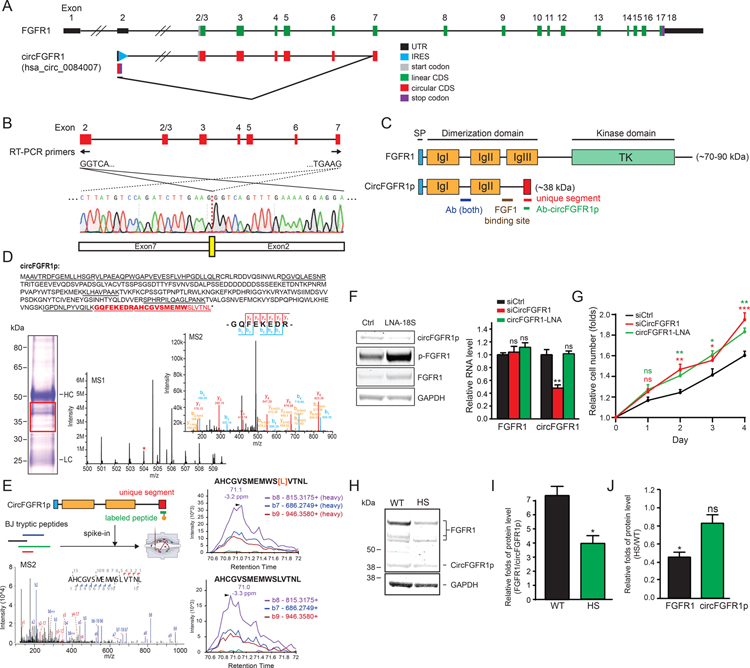
Comment in
-
Expanded regulation of circular RNA translation.Mol Cell. 2021 Oct 21;81(20):4111-4113. doi: 10.1016/j.molcel.2021.09.023. Mol Cell. 2021. PMID: 34686312
References
-
- Aitken CE, and Lorsch JR (2012). A mechanistic overview of translation initiation in eukaryotes. Nat Struct Mol Biol 19, 568–576. - PubMed
-
- Ashwal-Fluss R, Meyer M, Pamudurti NR, Ivanov A, Bartok O, Hanan M, Evantal N, Memczak S, Rajewsky N, and Kadener S (2014). circRNA biogenesis competes with pre-mRNA splicing. Mol Cell 56, 55–66. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
