

Spatial Cognition in Teleost Fish: Strategies and Mechanisms
- PMID: 34438729
- PMCID: PMC8388456
- DOI: 10.3390/ani11082271
Spatial Cognition in Teleost Fish: Strategies and Mechanisms
Abstract
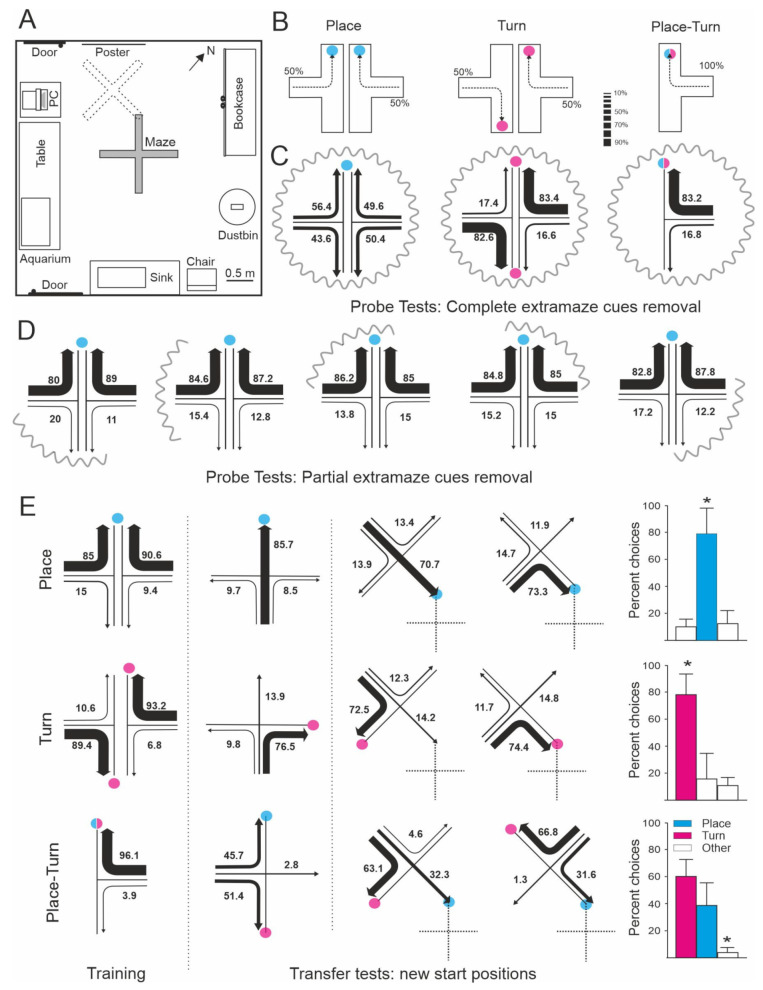
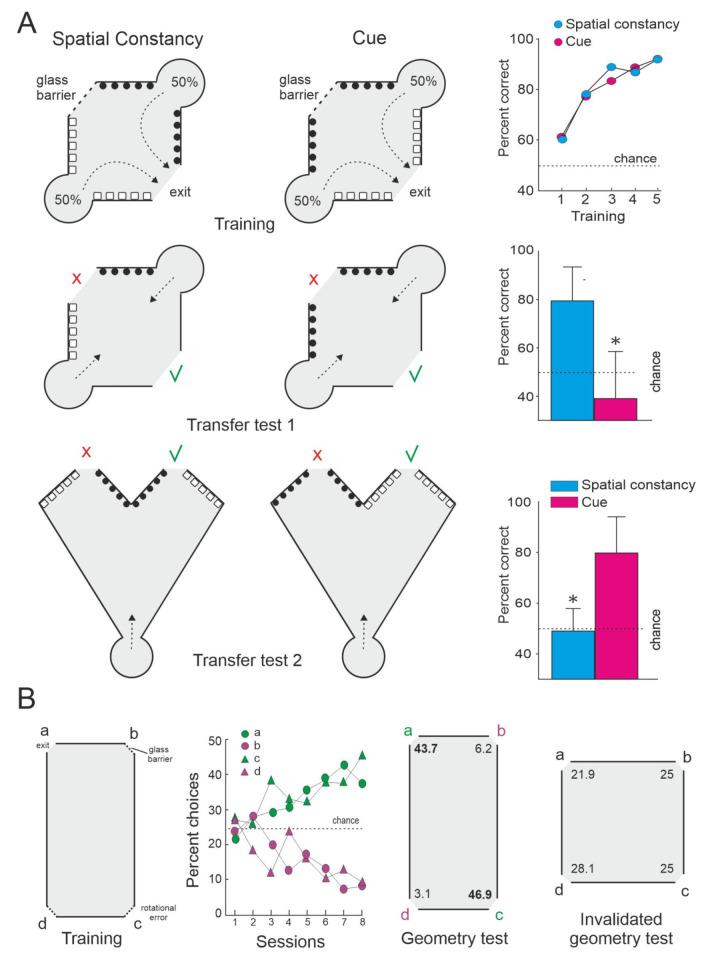
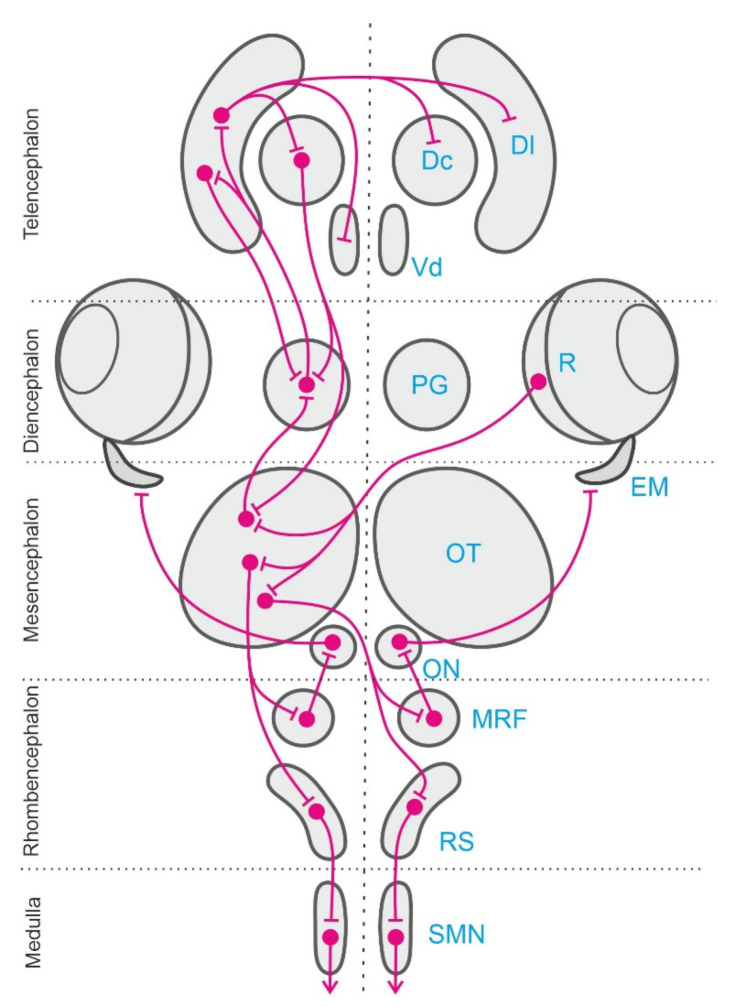
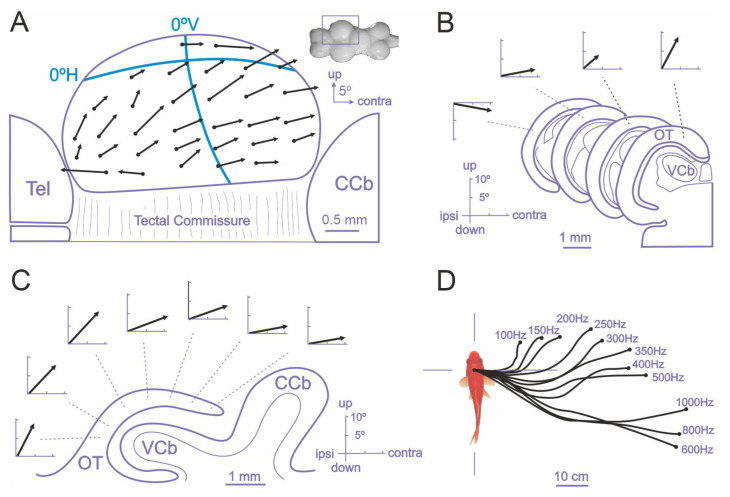
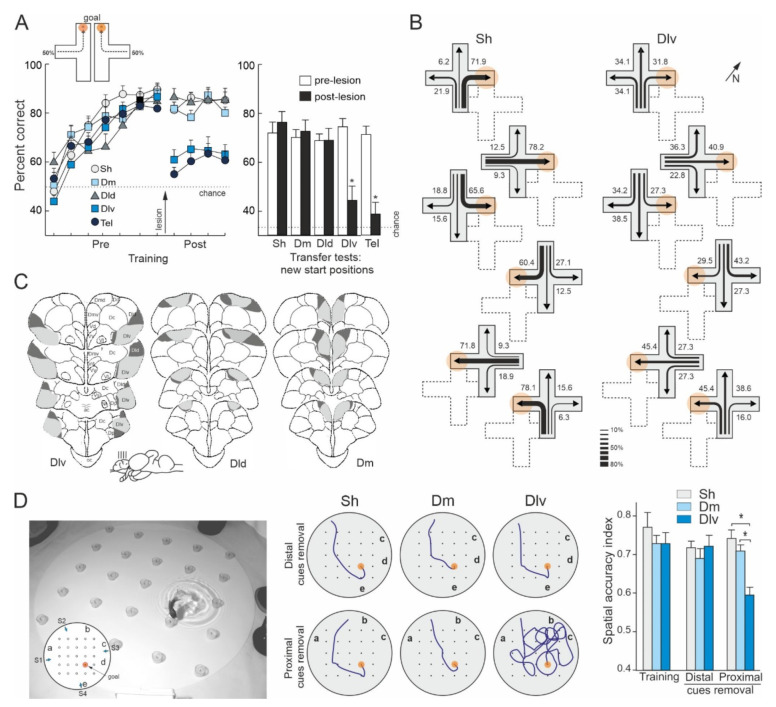
Teleost fish have been traditionally considered primitive vertebrates compared to mammals and birds in regard to brain complexity and behavioral functions. However, an increasing amount of evidence suggests that teleosts show advanced cognitive capabilities including spatial navigation skills that parallel those of land vertebrates. Teleost fish rely on a multiplicity of sensory cues and can use a variety of spatial strategies for navigation, ranging from relatively simple body-centered orientation responses to allocentric or "external world-centered" navigation, likely based on map-like relational memory representations of the environment. These distinct spatial strategies are based on separate brain mechanisms. For example, a crucial brain center for egocentric orientation in teleost fish is the optic tectum, which can be considered an essential hub in a wider brain network responsible for the generation of egocentrically referenced actions in space. In contrast, other brain centers, such as the dorsolateral telencephalic pallium of teleost fish, considered homologue to the hippocampal pallium of land vertebrates, seem to be crucial for allocentric navigation based on map-like spatial memory. Such hypothetical relational memory representations endow fish's spatial behavior with considerable navigational flexibility, allowing them, for example, to perform shortcuts and detours.
Keywords: hippocampal pallium; optic tectum; spatial navigation; spatial strategies; telencephalon; teleost fish; vertebrate brain evolution.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Dodson S. The ecological role of chemical stimuli for the zooplankton: Predator-avoidance behavior in Daphnia. Limnol. Oceanogr. 1988;33:1431–1439. doi: 10.4319/lo.1988.33.6_part_2.1431. - DOI
-
- Quinn T.P. Fishes. In: Papi F., editor. Animal Homing. Chapman & Hall; London, UK: 1992. pp. 145–211.
-
- Hallacher L.E. Relocation of original territories by displaced black-and-yellow rockfish, Sebastes chrysomelas, from Carmel Bay, California. Calif. Fish Game. 1984;7:158–162.
-
- Kroon F.J., de Graaf M., Liley N.R. Social organisation and competition for refuges and nest sites in Coryphopterus nicholsii (Gobiidae), a temperature protogynous reef fish. Environ. Biol. Fishes. 2000;57:401–411. doi: 10.1023/A:1007600728267. - DOI
-
- Matthews K.R. An experimental study of the habitat preferences and movement patterns of copper, quillback, and brown rockfishes (Sebastes spp.) Environ. Biol. Fishes. 1990;29:161–178. doi: 10.1007/BF00002217. - DOI
Publication types
LinkOut - more resources
Full Text Sources
