

The Interplay of Mitochondrial Oxidative Stress and Endoplasmic Reticulum Stress in Cardiovascular Fibrosis in Obese Rats
- PMID: 34439522
- PMCID: PMC8389298
- DOI: 10.3390/antiox10081274
The Interplay of Mitochondrial Oxidative Stress and Endoplasmic Reticulum Stress in Cardiovascular Fibrosis in Obese Rats
Abstract
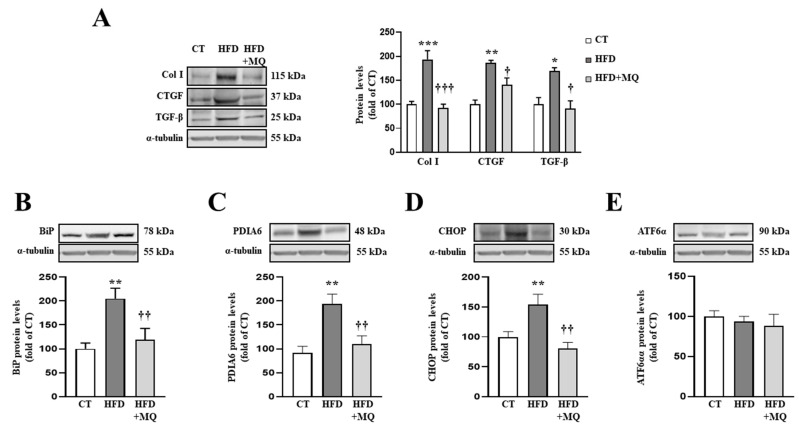
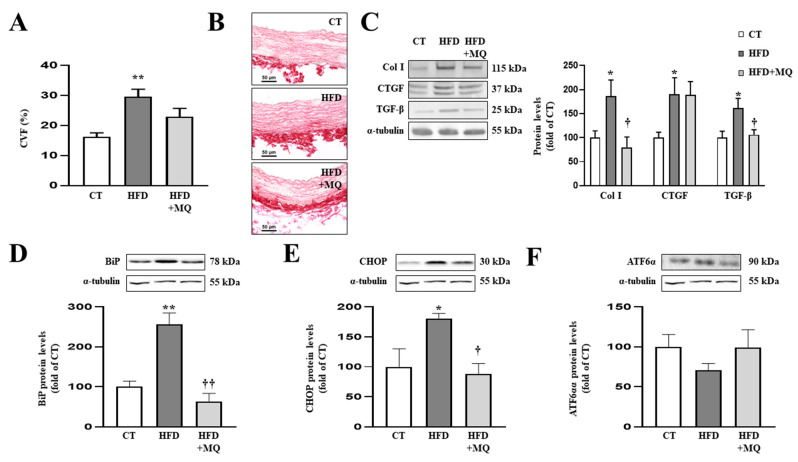
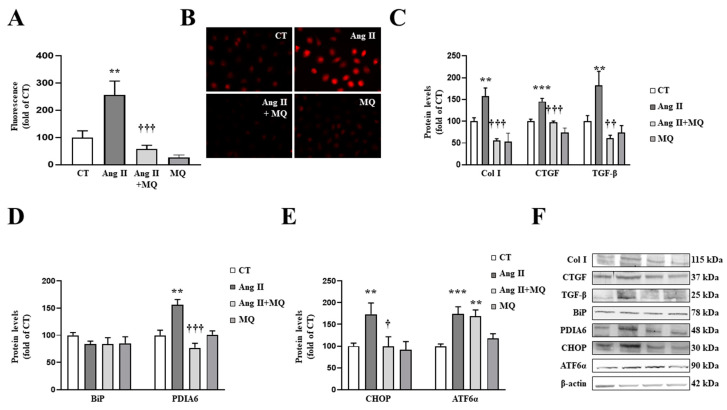
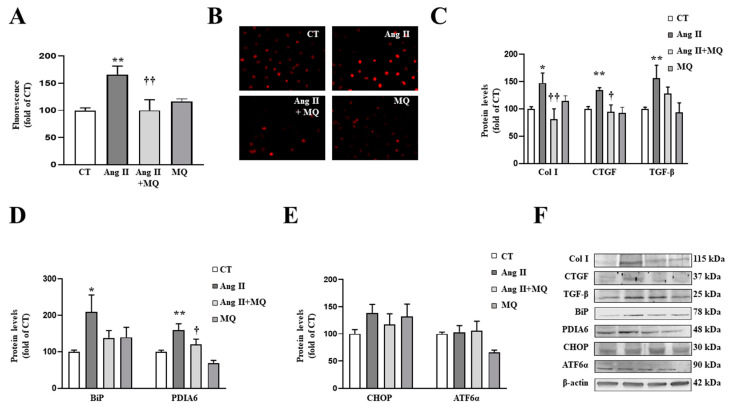
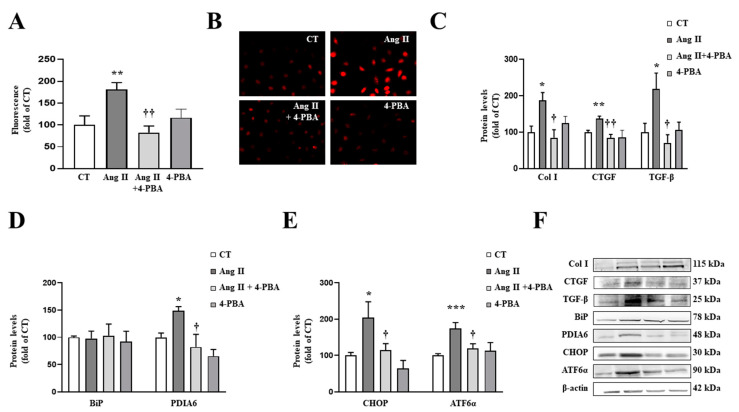
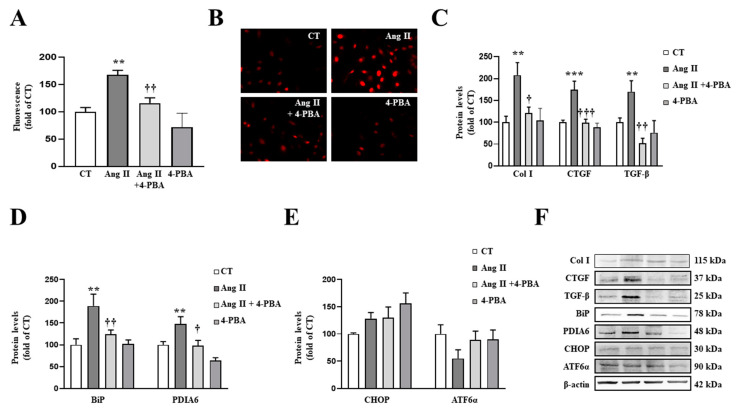
We have evaluated the role of mitochondrial oxidative stress and its association with endoplasmic reticulum (ER) stress activation in the progression of obesity-related cardiovascular fibrosis. MitoQ (200 µM) was orally administered for 7 weeks to male Wistar rats that were fed a high-fat diet (HFD, 35% fat) or a control diet (CT, 3.5% fat). Obese animals presented cardiovascular fibrosis accompanied by increased levels of extracellular matrix proteins and profibrotic mediators. These alterations were associated with ER stress activation characterized by enhanced levels (in heart and aorta vs. CT group, respectively) of immunoglobulin binding protein (BiP; 2.1-and 2.6-fold, respectively), protein disulfide-isomerase A6 (PDIA6; 1.9-fold) and CCAAT-enhancer-binding homologous protein (CHOP; 1.5- and 1.8-fold, respectively). MitoQ treatment was able to prevent (p < 0.05) these modifications at cardiac and aortic levels. MitoQ (5 nM) and the ER stress inhibitor, 4-phenyl butyric acid (4 µM), were able to block the prooxidant and profibrotic effects of angiotensin II (Ang II, 10-6 M) in cardiac and vascular cells. Therefore, the data show a crosstalk between mitochondrial oxidative stress and ER stress activation, which mediates the development of cardiovascular fibrosis in the context of obesity and in which Ang II can play a relevant role.
Keywords: cardiovascular fibrosis; endoplasmic reticulum stress; mitochondrial oxidative stress; obesity.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Martinez-Martinez E., Jurado-Lopez R., Valero-Munoz M., Bartolome M.V., Ballesteros S., Luaces M., Briones A.M., Lopez-Andres N., Miana M., Cachofeiro V. Leptin induces cardiac fibrosis through galectin-3, mTOR and oxidative stress: Potential role in obesity. J. Hypertens. 2014;32:1104–1114; discussion 1114. doi: 10.1097/HJH.0000000000000149. - DOI - PubMed
Grants and funding
- PI18/00257; PI15/00742; CIBERCV/Instituto de Salud Carlos III-Fondo Europeo de Desarrollo Regional (FEDER)
- CT42/18-CT43/18/Universidad Complutense de Madrid y Banco Santander
- FI19/00277/P-FIS
- 2017-T2/BMD-5523/contract from CAM (Atracción de talento)
- PEJD-2018-PRE/BMD-9171 and PEJ-2018-TL/BMD-11906/contract from CAM (Ayuda de empleo juvenil)
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
