

Long-Term Caffeine Intake Exerts Protective Effects on Intestinal Aging by Regulating Vitellogenesis and Mitochondrial Function in an Aged Caenorhabditis Elegans Model
- PMID: 34444677
- PMCID: PMC8398797
- DOI: 10.3390/nu13082517
Long-Term Caffeine Intake Exerts Protective Effects on Intestinal Aging by Regulating Vitellogenesis and Mitochondrial Function in an Aged Caenorhabditis Elegans Model
Abstract
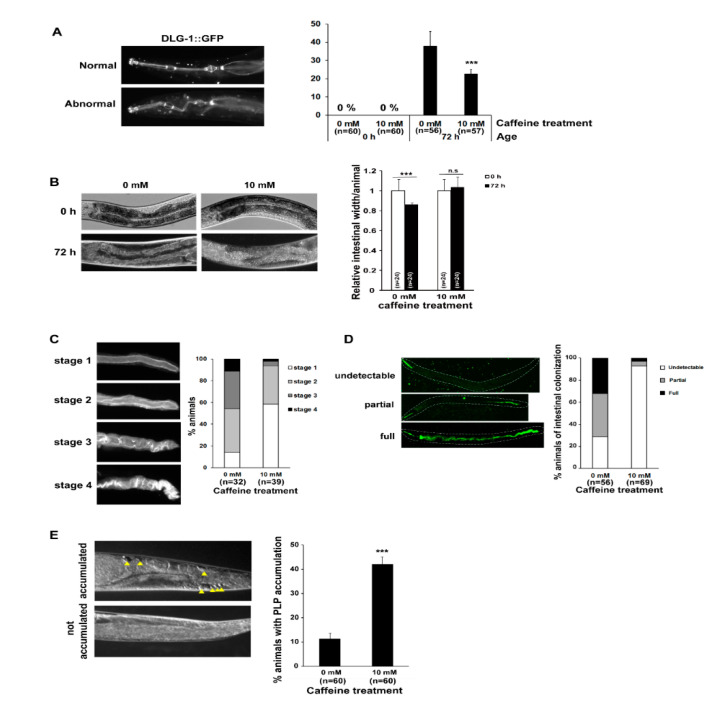
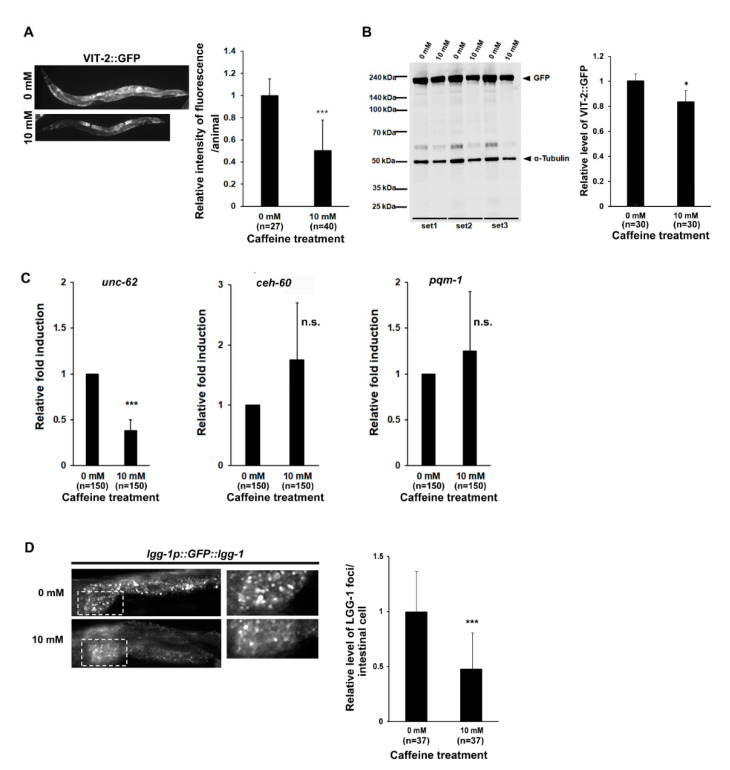
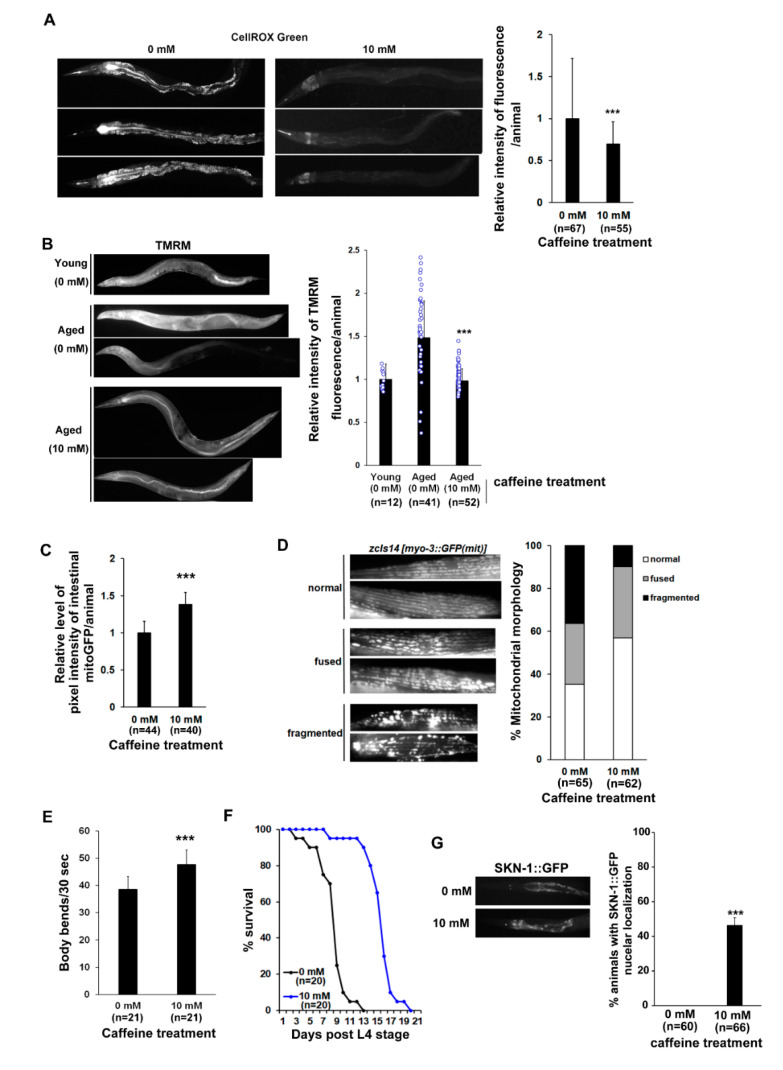
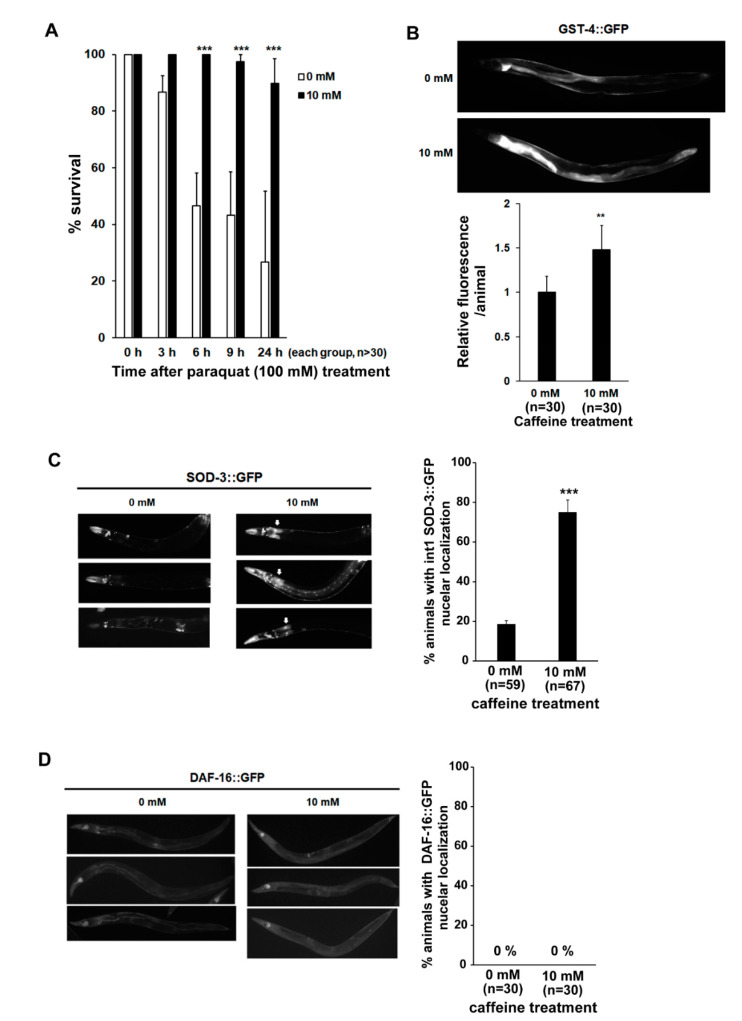
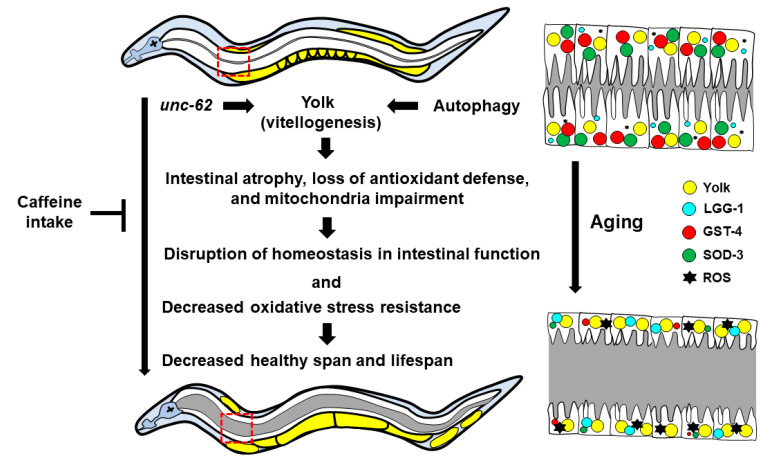
Caffeine, a methylxanthine derived from plants, is the most widely consumed ingredient in daily life. Therefore, it is necessary to investigate the effects of caffeine intake on essential biological activities. In this study, we attempted to determine the possible anti-aging effects of long-term caffeine intake in the intestine of an aged Caenorhabditis elegans model. We examined changes in intestinal integrity, production of vitellogenin (VIT), and mitochondrial function after caffeine intake. To evaluate intestinal aging, actin-5 (ACT-5) mislocalization, lumenal expansion, and intestinal colonization were examined after caffeine intake, and the levels of vitellogenesis as well as the mitochondrial activity were measured. We found that the long-term caffeine intake (10 mM) in the L4-stage worms at 25 °C for 3 days suppressed ACT-5 mislocalization. Furthermore, the level of autophagy, which is normally increased in aging animals, was significantly reduced in these animals, and their mitochondrial functions improved after caffeine intake. In addition, the caffeine-ingesting aging animals showed high resistance to oxidative stress and increased the expression of antioxidant proteins. Taken together, these findings reveal that caffeine may be a potential anti-aging agent that can suppress intestinal atrophy during the progression of intestinal aging.
Keywords: Caenorhabditis elegans; anti-aging; caffeine; intestinal aging; mitochondrial function; oxidative stress response; vitellogenesis.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Du X., Guan Y., Huang Q., Lv M., He X., Yan Y., Hayashi S., Fang C., Wang X., Sheng J. Low concentrations of caffeine and its analogs extend the lifespan of Caenorhabditis elegans by modulating IGF-1-Like pathway. Front. Aging Neurosci. 2018;10:211. doi: 10.3389/fnagi.2018.00211. - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
