

Moisture modulates soil reservoirs of active DNA and RNA viruses
- PMID: 34446837
- PMCID: PMC8390657
- DOI: 10.1038/s42003-021-02514-2
Moisture modulates soil reservoirs of active DNA and RNA viruses
Abstract
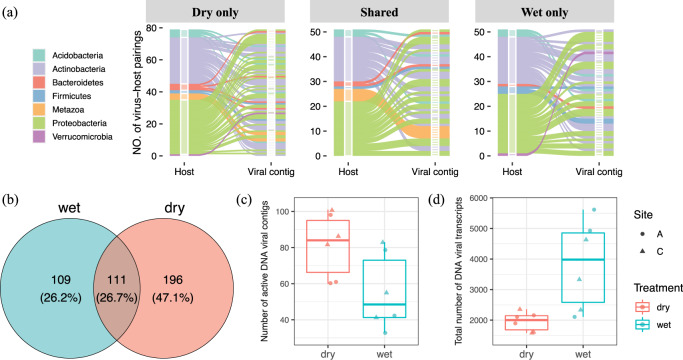
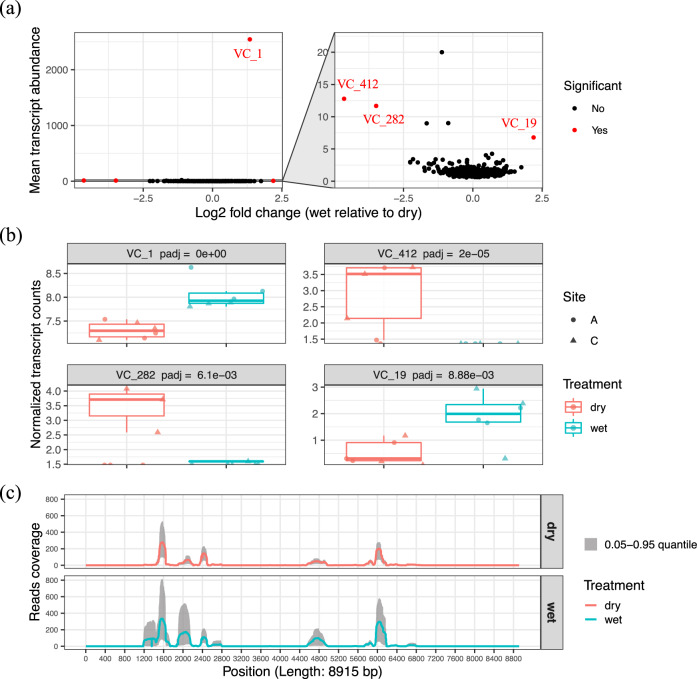
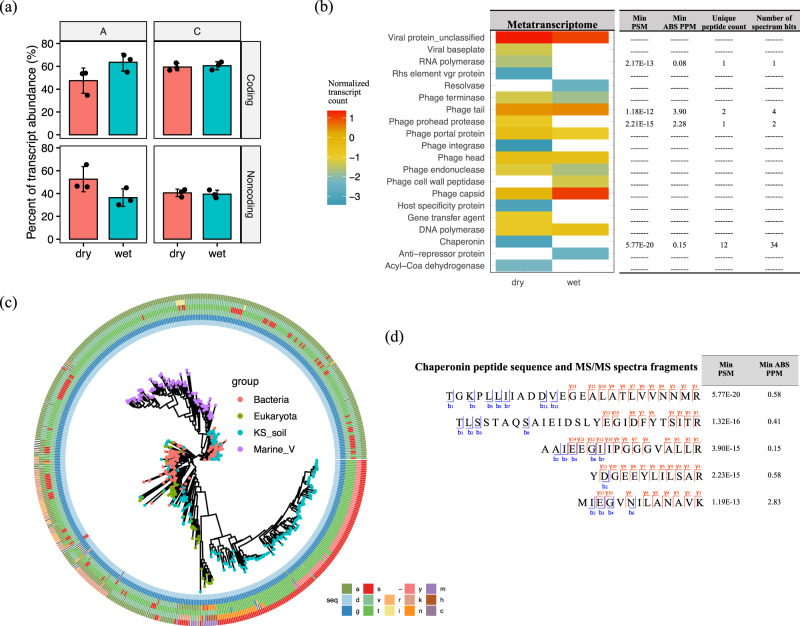
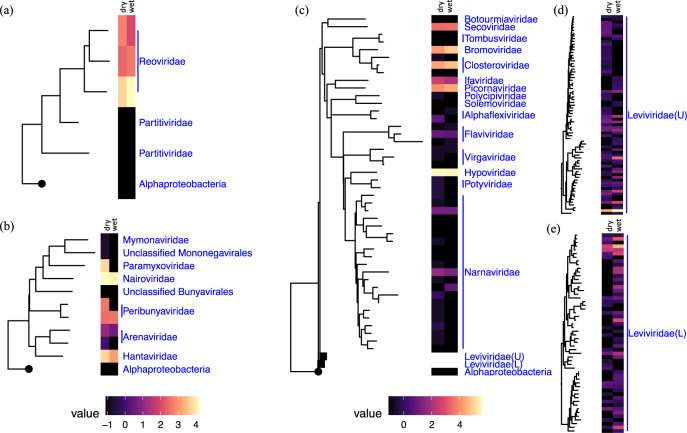
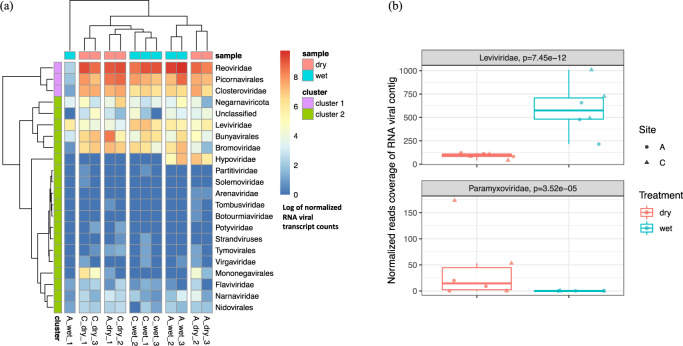
Soil is known to harbor viruses, but the majority are uncharacterized and their responses to environmental changes are unknown. Here, we used a multi-omics approach (metagenomics, metatranscriptomics and metaproteomics) to detect active DNA viruses and RNA viruses in a native prairie soil and to determine their responses to extremes in soil moisture. The majority of transcribed DNA viruses were bacteriophage, but some were assigned to eukaryotic hosts, mainly insects. We also demonstrated that higher soil moisture increased transcription of a subset of DNA viruses. Metaproteome data validated that the specific viral transcripts were translated into proteins, including chaperonins known to be essential for virion replication and assembly. The soil viral chaperonins were phylogenetically distinct from previously described marine viral chaperonins. The soil also had a high abundance of RNA viruses, with highest representation of Reoviridae. Leviviridae were the most diverse RNA viruses in the samples, with higher amounts in wet soil. This study demonstrates that extreme shifts in soil moisture have dramatic impacts on the composition, activity and potential functions of both DNA and RNA soil viruses.
© 2021. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
