

Drosophila re-zero their path integrator at the center of a fictive food patch
- PMID: 34450090
- PMCID: PMC8551043
- DOI: 10.1016/j.cub.2021.08.006
Drosophila re-zero their path integrator at the center of a fictive food patch
Abstract
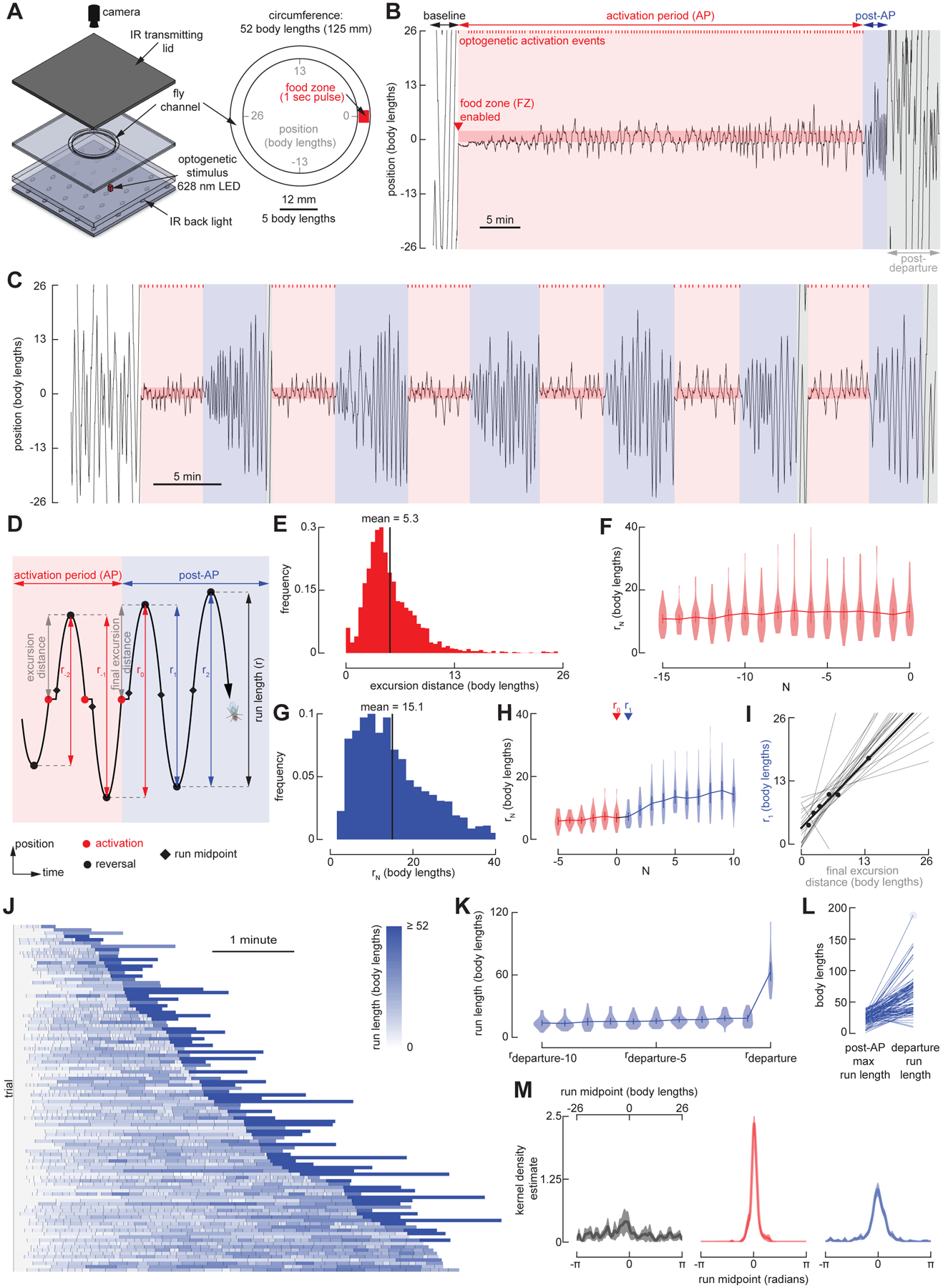
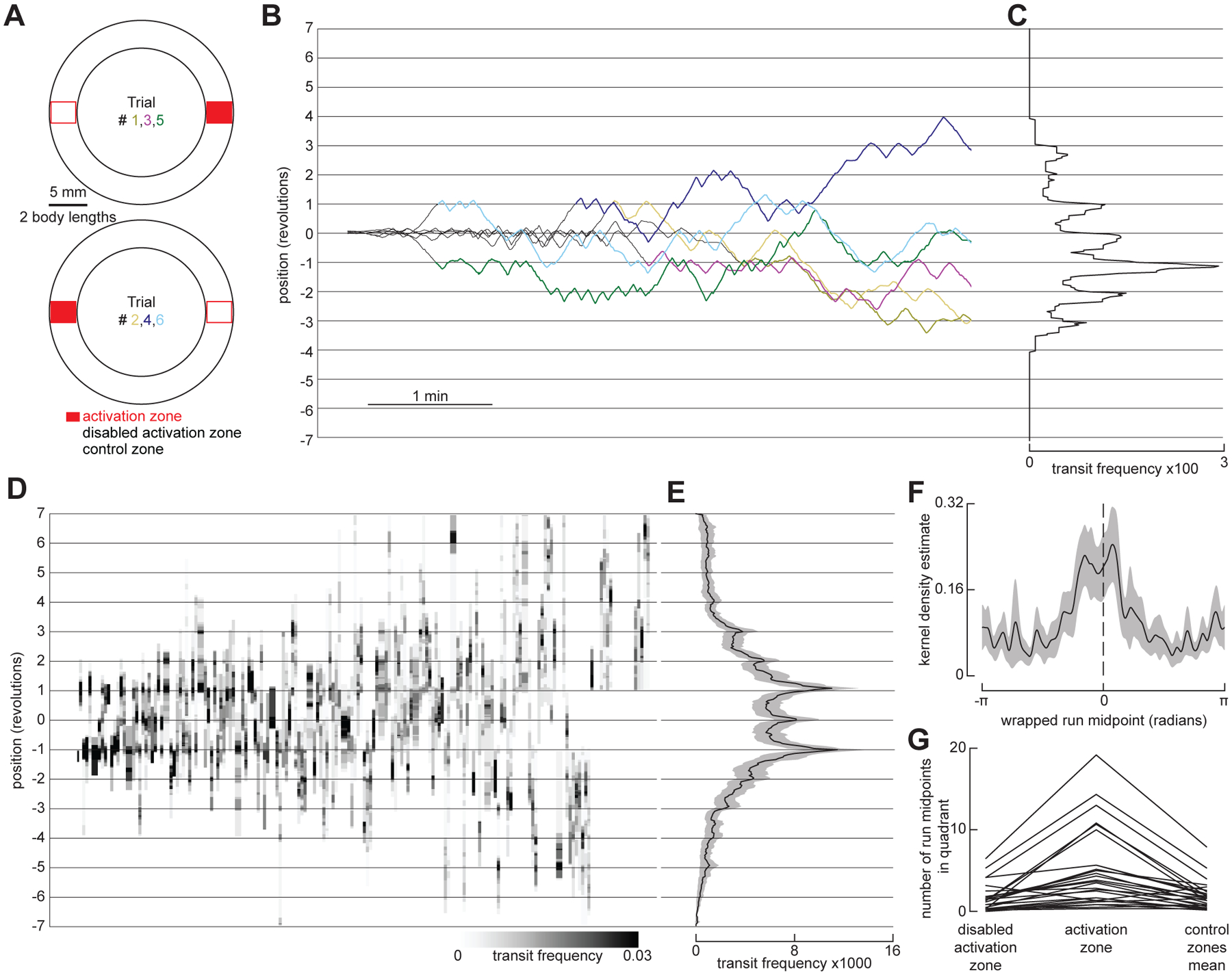
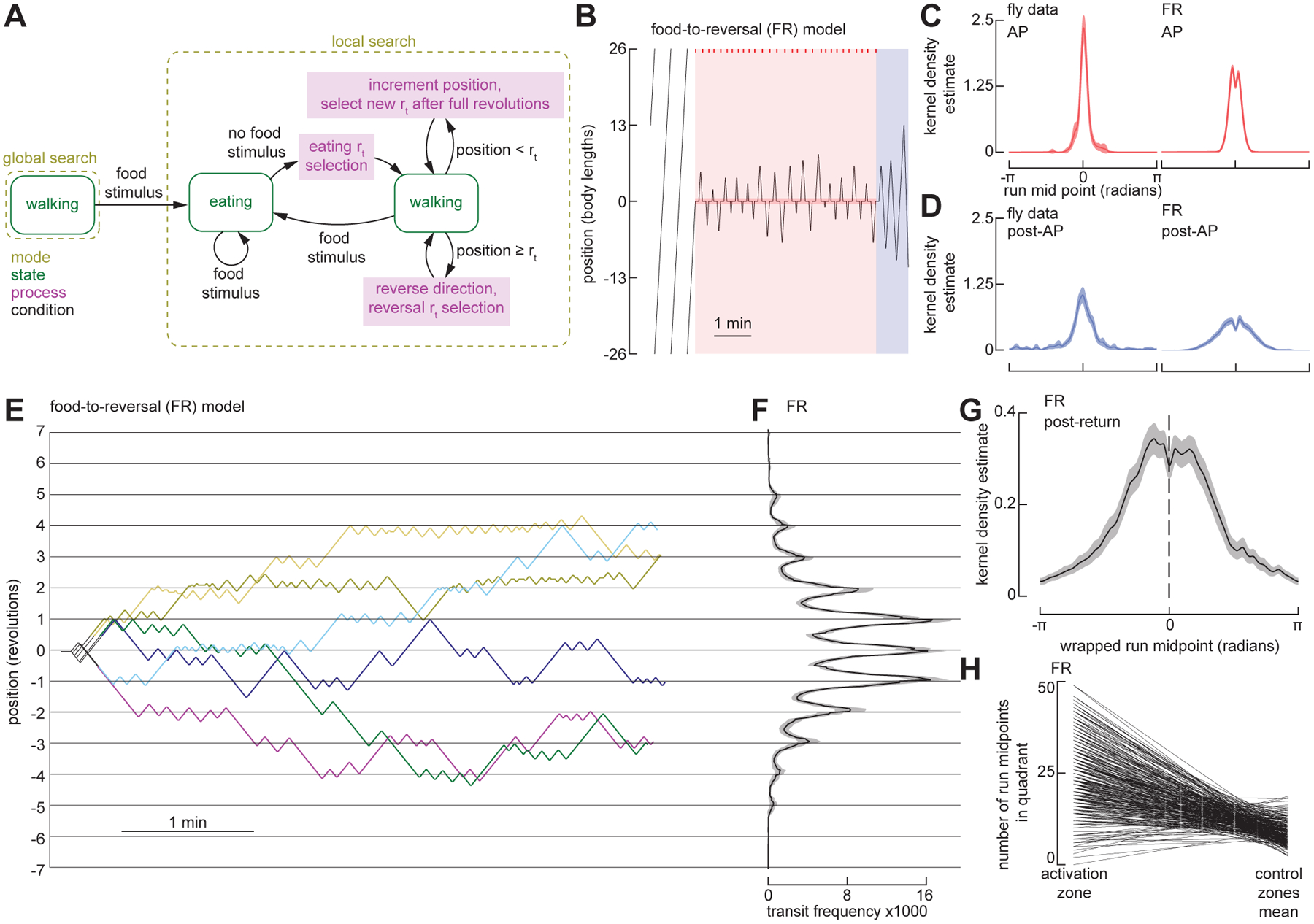
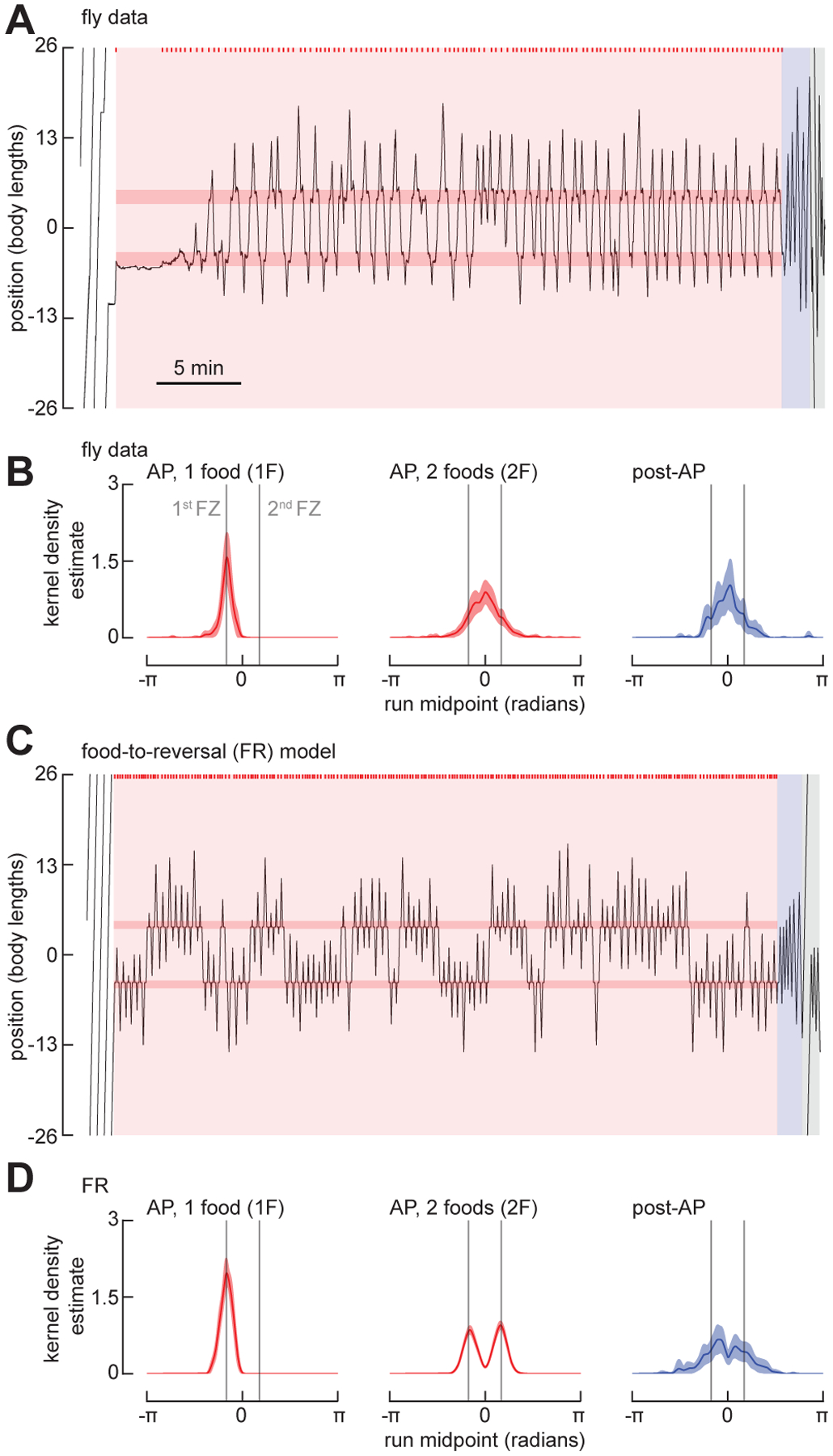
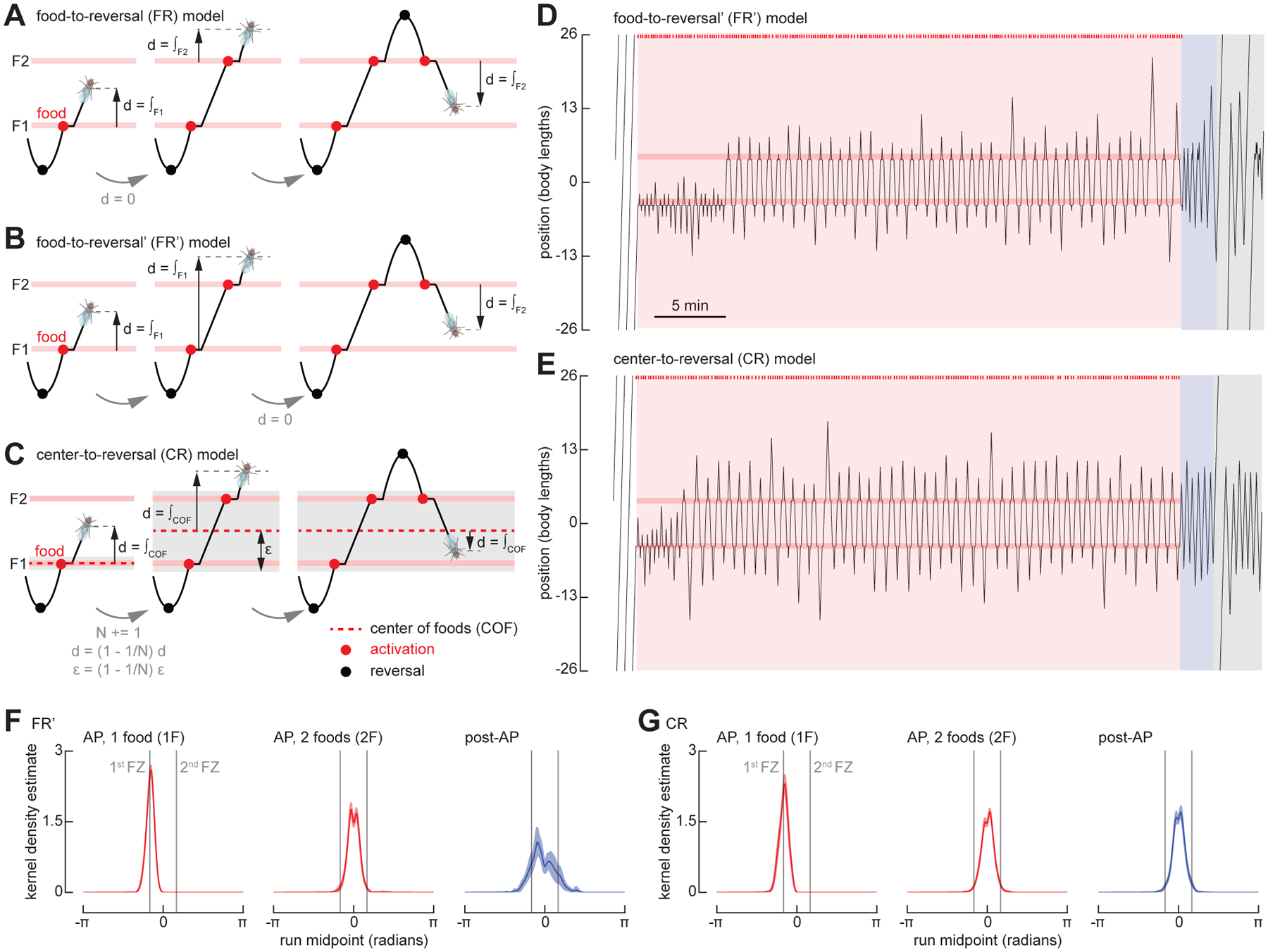
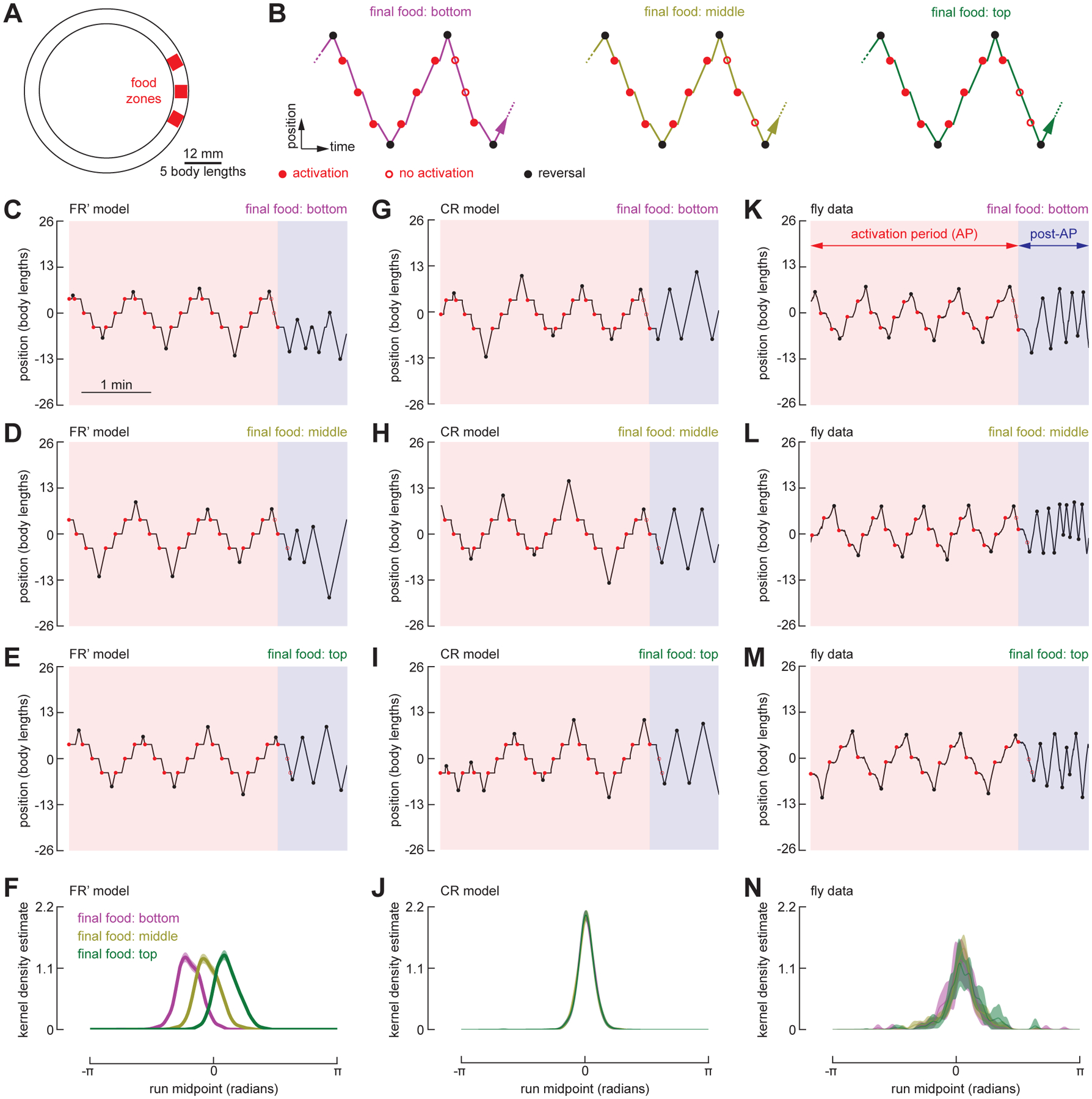
The ability to keep track of one's location in space is a critical behavior for animals navigating to and from a salient location, and its computational basis is now beginning to be unraveled. Here, we tracked flies in a ring-shaped channel as they executed bouts of search triggered by optogenetic activation of sugar receptors. Unlike experiments in open field arenas, which produce highly tortuous search trajectories, our geometrically constrained paradigm enabled us to monitor flies' decisions to move toward or away from the fictive food. Our results suggest that flies use path integration to remember the location of a food site even after it has disappeared, and flies can remember the location of a former food site even after walking around the arena one or more times. To determine the behavioral algorithms underlying Drosophila search, we developed multiple state transition models and found that flies likely accomplish path integration by combining odometry and compass navigation to keep track of their position relative to the fictive food. Our results indicate that whereas flies re-zero their path integrator at food when only one feeding site is present, they adjust their path integrator to a central location between sites when experiencing food at two or more locations. Together, this work provides a simple experimental paradigm and theoretical framework to advance investigations of the neural basis of path integration.
Keywords: Drosophila; odometry; path integration; place memory; state-dependent models.
Copyright © 2021 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
Comment in
-
Fly navigation: Yet another ring.Curr Biol. 2021 Oct 25;31(20):R1381-R1383. doi: 10.1016/j.cub.2021.09.009. Curr Biol. 2021. PMID: 34699800
References
-
- Pyke GH (1984). Optimal foraging theory. Annu. Rev. Ecol. Syst 15, 523–575.
-
- Whishaw IQ, and Brooks BL (1999). Calibrating space: exploration is important for allothetic and idiothetic navigation. Hippocampus 9, 659–667. - PubMed
-
- Wolf H (2011). Odometry and insect navigation. J. Exp. Biol 214, 1629–1641. - PubMed
-
- Bühlmann C, Cheng K, and Wehner R (2011). Vector-based and landmark-guided navigation in desert ants inhabiting landmark-free and landmark-rich environments. J. Exp. Biol 214, 2845–2853. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
