

Environmental vulnerability of the global ocean epipelagic plankton community interactome
- PMID: 34452910
- PMCID: PMC8397264
- DOI: 10.1126/sciadv.abg1921
Environmental vulnerability of the global ocean epipelagic plankton community interactome
Abstract
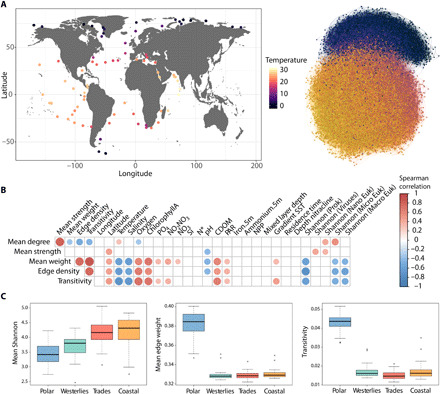
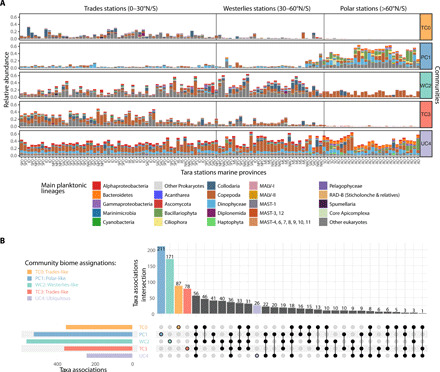
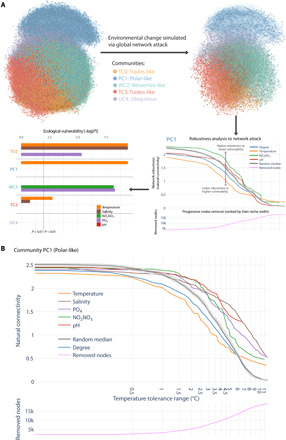
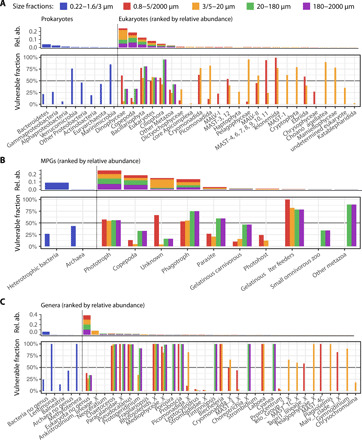
Marine plankton form complex communities of interacting organisms at the base of the food web, which sustain oceanic biogeochemical cycles and help regulate climate. Although global surveys are starting to reveal ecological drivers underlying planktonic community structure and predicted climate change responses, it is unclear how community-scale species interactions will be affected by climate change. Here, we leveraged Tara Oceans sampling to infer a global ocean cross-domain plankton co-occurrence network-the community interactome-and used niche modeling to assess its vulnerabilities to environmental change. Globally, this revealed a plankton interactome self-organized latitudinally into marine biomes (Trades, Westerlies, Polar) and more connected poleward. Integrated niche modeling revealed biome-specific community interactome responses to environmental change and forecasted the most affected lineages for each community. These results provide baseline approaches to assess community structure and organismal interactions under climate scenarios while identifying plausible plankton bioindicators for ocean monitoring of climate change.
Copyright © 2021 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works. Distributed under a Creative Commons Attribution License 4.0 (CC BY).
Figures
References
-
- Field C. B., Behrenfeld M. J., Randerson J. T., Falkowski P., Primary production of the biosphere: Integrating terrestrial and oceanic components. Science 281, 237–240 (1998). - PubMed
-
- Gregory A. C., Zayed A. A., Conceição-Neto N., Temperton B., Bolduc B., Alberti A., Ardyna M., Arkhipova K., Carmichael M., Cruaud C., Dimier C., Domínguez-Huerta G., Ferland J., Kandels S., Liu Y., Marec C., Pesant S., Picheral M., Pisarev S., Poulain J., Tremblay J.-É., Vik D.; Tara Oceans Coordinators, Babin M., Bowler C., Culley A. I., de Vargas C., Dutilh B. E., Iudicone D., Karp-Boss L., Roux S., Sunagawa S., Wincker P., Sullivan M. B., Marine DNA viral macro- and microdiversity from pole to pole. Cell 177, 1109–1123.e14 (2019). - PMC - PubMed
-
- Ibarbalz F. M., Henry N., Brandão M. C., Martini S., Busseni G., Byrne H., Coelho L. P., Endo H., Gasol J. M., Gregory A. C., Mahé F., Rigonato J., Royo-Llonch M., Salazar G., Sanz-Sáez I., Scalco E., Soviadan D., Zayed A. A., Zingone A., Labadie K., Ferland J., Marec C., Kandels S., Picheral M., Dimier C., Poulain J., Pisarev S., Carmichael M., Pesant S.; Tara Oceans Coordinators, Babin M., Boss E., Iudicone D., Jaillon O., Acinas S. G., Ogata H., Pelletier E., Stemmann L., Sullivan M. B., Sunagawa S., Bopp L., de Vargas C., Karp-Boss L., Wincker P., Lombard F., Bowler C., Zinger L., Global trends in marine plankton diversity across kingdoms of life. Cell 179, 1084–1097.e21 (2019). - PMC - PubMed
-
- Salazar G., Paoli L., Alberti A., Huerta-Cepas J., Ruscheweyh H.-J., Cuenca M., Field C. M., Coelho L. P., Cruaud C., Engelen S., Gregory A. C., Labadie K., Marec C., Pelletier E., Royo-Llonch M., Roux S., Sánchez P., Uehara H., Zayed A. A., Zeller G., Carmichael M., Dimier C., Ferland J., Kandels S., Picheral M., Pisarev S., Poulain J.; Tara Oceans Coordinators, Acinas S. G., Babin M., Bork P., Bowler C., de Vargas C., Guidi L., Hingamp P., Iudicone D., Karp-Boss L., Karsenti E., Ogata H., Pesant S., Speich S., Sullivan M. B., Wincker P., Sunagawa S., Gene expression changes and community turnover differentially shape the global ocean metatranscriptome. Cell 179, 1068–1083.e21 (2019). - PMC - PubMed
-
- Stocker R., Marine microbes see a sea of gradients. Science 338, 628–633 (2012). - PubMed
LinkOut - more resources
Full Text Sources
