

Structural basis for the activation of the DEAD-box RNA helicase DbpA by the nascent ribosome
- PMID: 34453003
- PMCID: PMC8536315
- DOI: 10.1073/pnas.2105961118
Structural basis for the activation of the DEAD-box RNA helicase DbpA by the nascent ribosome
Abstract
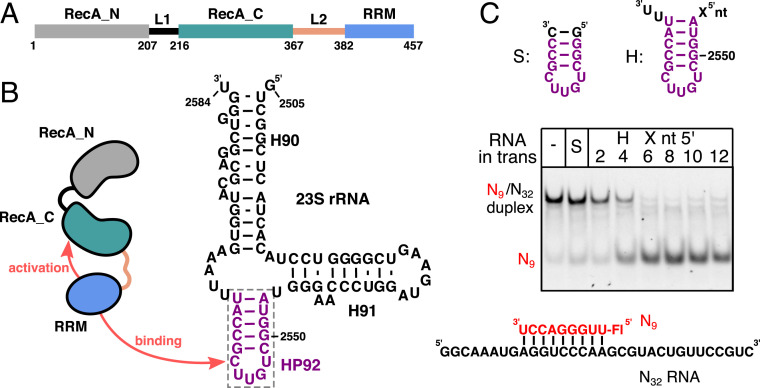
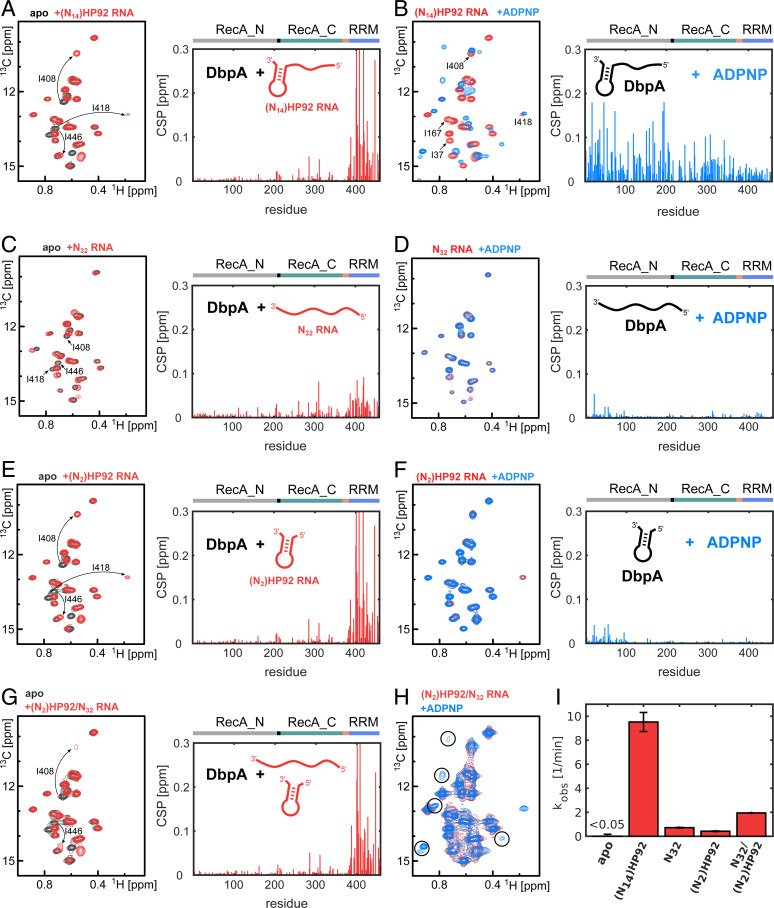
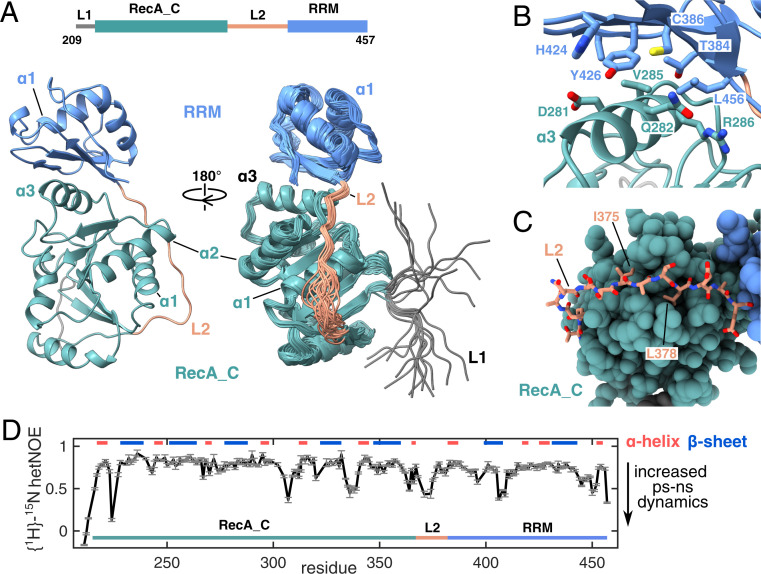
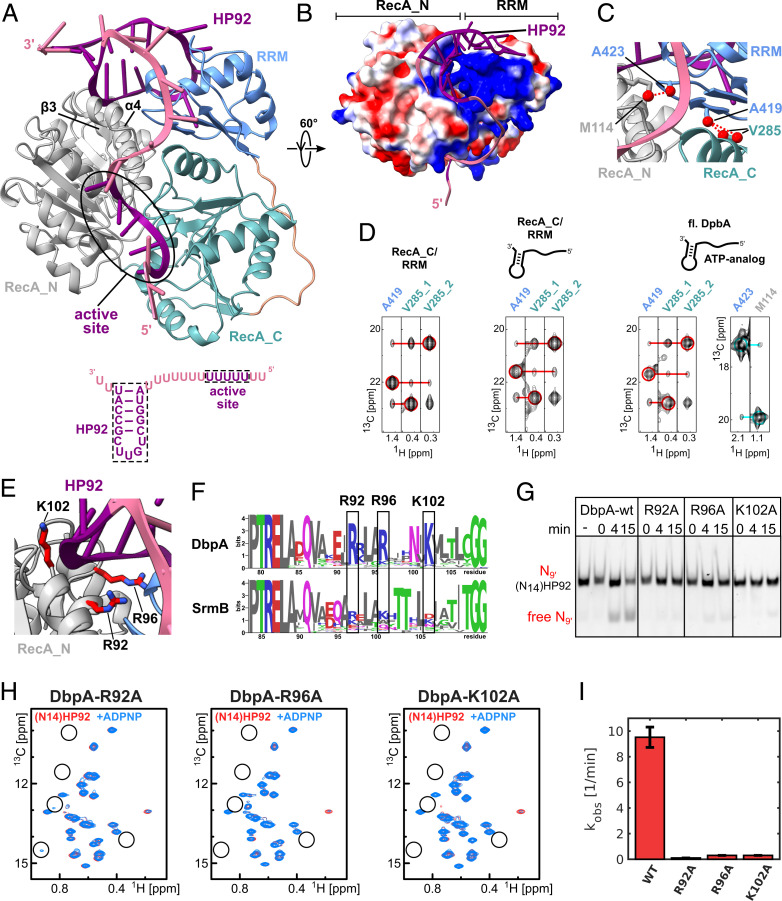
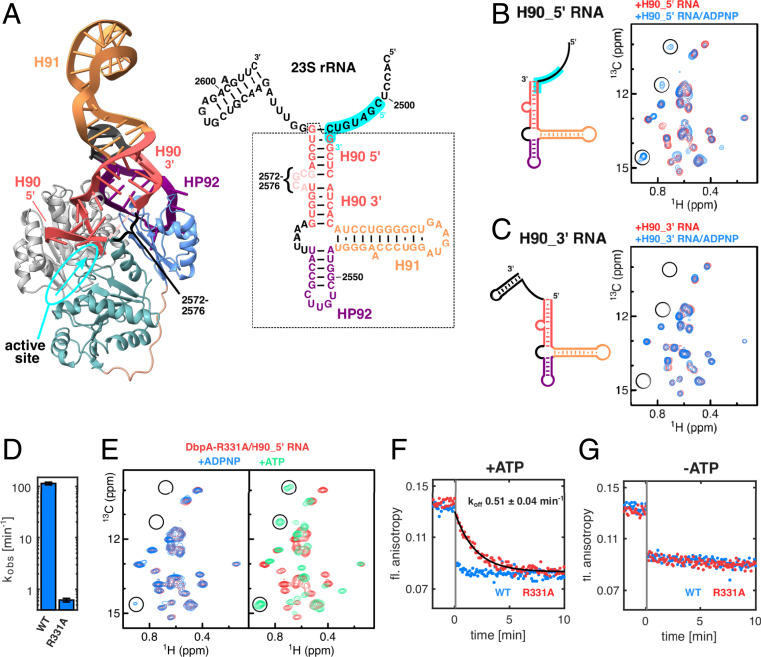
The adenosine triphosphate (ATP)-dependent DEAD-box RNA helicase DbpA from Escherichia coli functions in ribosome biogenesis. DbpA is targeted to the nascent 50S subunit by an ancillary, carboxyl-terminal RNA recognition motif (RRM) that specifically binds to hairpin 92 (HP92) of the 23S ribosomal RNA (rRNA). The interaction between HP92 and the RRM is required for the helicase activity of the RecA-like core domains of DbpA. Here, we elucidate the structural basis by which DbpA activity is endorsed when the enzyme interacts with the maturing ribosome. We used nuclear magnetic resonance (NMR) spectroscopy to show that the RRM and the carboxyl-terminal RecA-like domain tightly interact. This orients HP92 such that this RNA hairpin can form electrostatic interactions with a positively charged patch in the N-terminal RecA-like domain. Consequently, the enzyme can stably adopt the catalytically important, closed conformation. The substrate binding mode in this complex reveals that a region 5' to helix 90 in the maturing ribosome is specifically targeted by DbpA. Finally, our results indicate that the ribosome maturation defects induced by a dominant negative DbpA mutation are caused by a delayed dissociation of DbpA from the nascent ribosome. Taken together, our findings provide unique insights into the important regulatory mechanism that modulates the activity of DbpA.
Keywords: DEAD-box helicase; NMR spectroscopy; enzyme regulation; molecular mechanism; ribosome assembly.
Copyright © 2021 the Author(s). Published by PNAS.
Conflict of interest statement
The authors declare no competing interest.
Figures
Similar articles
-
The DbpA catalytic core unwinds double-helix substrates by directly loading on them.RNA. 2016 Mar;22(3):408-15. doi: 10.1261/rna.052928.115. Epub 2016 Jan 11. RNA. 2016. PMID: 26755693 Free PMC article.
-
Ribosome assembly in Escherichia coli strains lacking the RNA helicase DeaD/CsdA or DbpA.FEBS J. 2008 Aug;275(15):3772-82. doi: 10.1111/j.1742-4658.2008.06523.x. Epub 2008 Jun 28. FEBS J. 2008. PMID: 18565105
-
Assignment of the Ile, Leu, Val, Met and Ala methyl group resonances of the DEAD-box RNA helicase DbpA from E. coli.Biomol NMR Assign. 2021 Apr;15(1):121-128. doi: 10.1007/s12104-020-09994-z. Epub 2020 Dec 4. Biomol NMR Assign. 2021. PMID: 33277687 Free PMC article.
-
Assembly of bacterial ribosomes.Annu Rev Biochem. 2011;80:501-26. doi: 10.1146/annurev-biochem-062608-160432. Annu Rev Biochem. 2011. PMID: 21529161 Review.
-
DEAD Box RNA Helicases: Biochemical Properties, Role in RNA Processing and Ribosome Biogenesis.Cell Biochem Biophys. 2024 Jun;82(2):427-434. doi: 10.1007/s12013-024-01240-w. Epub 2024 Mar 2. Cell Biochem Biophys. 2024. PMID: 38430409 Review.
Cited by
-
Disassembly of unstable RNA structures by an E. coli DEAD-box chaperone accelerates ribosome assembly.Nucleic Acids Res. 2025 Feb 8;53(4):gkaf104. doi: 10.1093/nar/gkaf104. Nucleic Acids Res. 2025. PMID: 39988318 Free PMC article.
-
Structural basis for RNA-duplex unwinding by the DEAD-box helicase DbpA.RNA. 2023 Sep;29(9):1339-1354. doi: 10.1261/rna.079582.123. Epub 2023 May 23. RNA. 2023. PMID: 37221012 Free PMC article.
-
YjgA plays dual roles in enhancing PTC maturation.Nucleic Acids Res. 2024 Jul 22;52(13):7947-7960. doi: 10.1093/nar/gkae469. Nucleic Acids Res. 2024. PMID: 38842932 Free PMC article.
-
Sequence-specific remodeling of a topologically complex RNP substrate by Spb4.Nat Struct Mol Biol. 2022 Dec;29(12):1228-1238. doi: 10.1038/s41594-022-00874-9. Epub 2022 Dec 8. Nat Struct Mol Biol. 2022. PMID: 36482249 Free PMC article.
-
Enteroviral 2C protein is an RNA-stimulated ATPase and uses a two-step mechanism for binding to RNA and ATP.Nucleic Acids Res. 2022 Nov 11;50(20):11775-11798. doi: 10.1093/nar/gkac1054. Nucleic Acids Res. 2022. PMID: 36399514 Free PMC article.
References
-
- Linder P., Jankowsky E., From unwinding to clamping - The DEAD box RNA helicase family. Nat. Rev. Mol. Cell Biol. 12, 505–516 (2011). - PubMed
-
- Sengoku T., Nureki O., Nakamura A., Kobayashi S., Yokoyama S., Structural basis for RNA unwinding by the DEAD-box protein Drosophila Vasa. Cell 125, 287–300 (2006). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
