

Why do insects evolve immune priming? A search for crossroads
- PMID: 34453994
- PMCID: PMC7614680
- DOI: 10.1016/j.dci.2021.104246
Why do insects evolve immune priming? A search for crossroads
Abstract
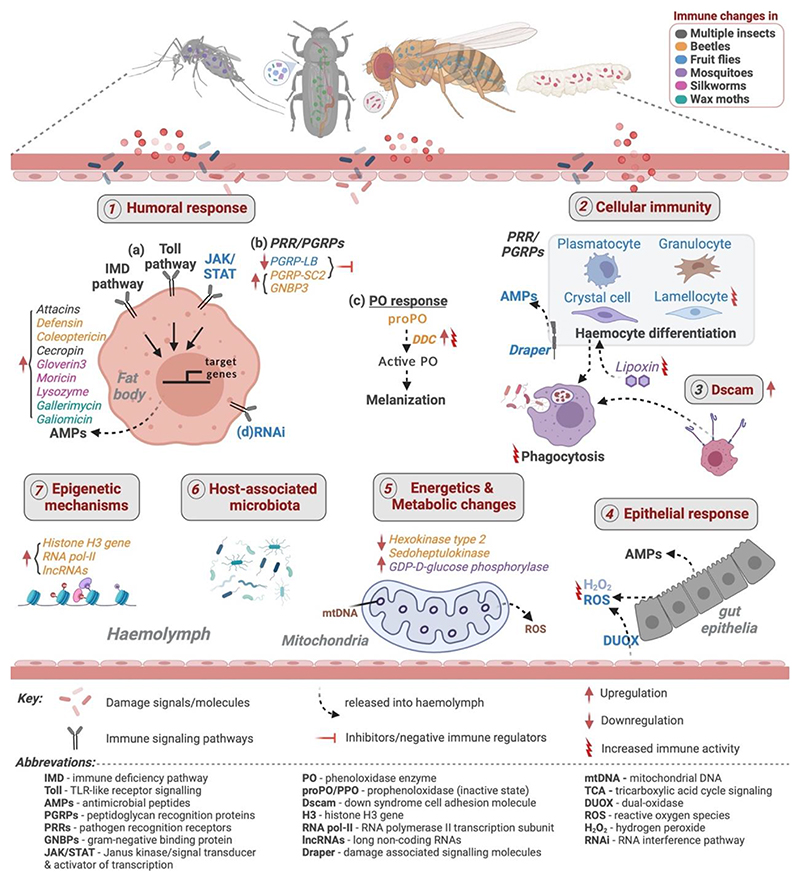
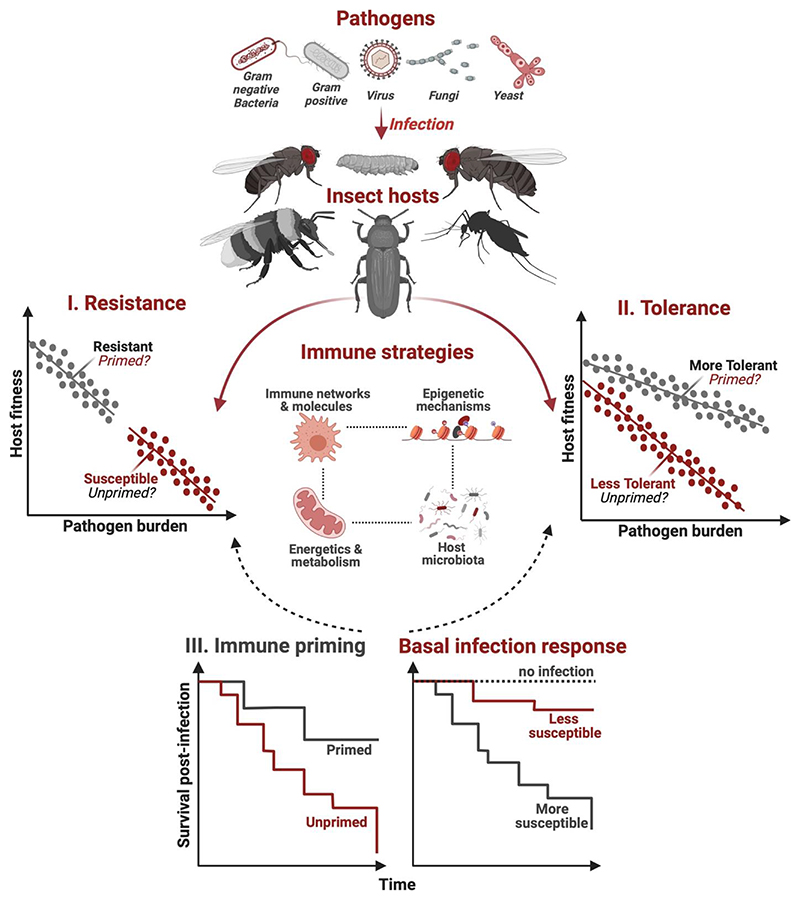
Until recently, it was assumed that insects lack immune memory since they do not have vertebrate-like specialized memory cells. Therefore, their most well studied evolutionary response against pathogens was increased basal immunity. However, growing evidence suggests that many insects also exhibit a form of immune memory (immune priming), where prior exposure to a low dose of infection confers protection against subsequent infection by the same pathogen that acts both within and across generations. Most strikingly, they can rapidly evolve as a highly parallel and mutually exclusive strategy from basal immunity, under different selective conditions and with divergent evolutionary trade-offs. However, the relative importance of priming as an optimal immune strategy also has contradictions, primarily because supporting mechanisms are still unclear. In this review, we adopt a comparative approach to highlight several emerging evolutionary, ecological and mechanistic features of priming vs basal immune responses that warrant immediate attention for future research.
Keywords: Coinfections; Cost of immunity; Immune priming; Resistance; Specificity; Tolerance.
Copyright © 2021 Elsevier Ltd. All rights reserved.
Conflict of interest statement
We have no competing interests
Figures
Similar articles
-
Experimental evolution of insect immune memory versus pathogen resistance.Proc Biol Sci. 2017 Dec 20;284(1869):20171583. doi: 10.1098/rspb.2017.1583. Proc Biol Sci. 2017. PMID: 29237849 Free PMC article.
-
Current understanding of immune priming phenomena in insects.J Invertebr Pathol. 2021 Oct;185:107656. doi: 10.1016/j.jip.2021.107656. Epub 2021 Aug 28. J Invertebr Pathol. 2021. PMID: 34464656 Review.
-
Immune priming specificity within and across generations reveals the range of pathogens affecting evolution of immunity in an insect.J Anim Ecol. 2018 Mar;87(2):448-463. doi: 10.1111/1365-2656.12661. Epub 2017 Apr 11. J Anim Ecol. 2018. PMID: 28239855
-
Divergent immune priming responses across flour beetle life stages and populations.Ecol Evol. 2016 Oct 9;6(21):7847-7855. doi: 10.1002/ece3.2532. eCollection 2016 Nov. Ecol Evol. 2016. PMID: 30128134 Free PMC article.
-
Immune priming: the secret weapon of the insect world.Virulence. 2020 Dec;11(1):238-246. doi: 10.1080/21505594.2020.1731137. Virulence. 2020. PMID: 32079502 Free PMC article. Review.
Cited by
-
The costs of transgenerational immune priming for homologous and heterologous infections with different serotypes of dengue virus in Aedes aegypti mosquitoes.Front Immunol. 2023 Dec 18;14:1286831. doi: 10.3389/fimmu.2023.1286831. eCollection 2023. Front Immunol. 2023. PMID: 38170025 Free PMC article.
-
Influences of Microbial Symbionts on Chemoreception of Their Insect Hosts.Insects. 2023 Jul 14;14(7):638. doi: 10.3390/insects14070638. Insects. 2023. PMID: 37504644 Free PMC article. Review.
-
Drosophila melanogaster as a model to study innate immune memory.Front Microbiol. 2022 Oct 20;13:991678. doi: 10.3389/fmicb.2022.991678. eCollection 2022. Front Microbiol. 2022. PMID: 36338030 Free PMC article. Review.
-
Sociality and disease: behavioral perspectives in ecological and evolutionary immunology.Behav Ecol Sociobiol. 2022;76(7):98. doi: 10.1007/s00265-022-03203-8. Epub 2022 Jul 8. Behav Ecol Sociobiol. 2022. PMID: 35821673 Free PMC article. No abstract available.
-
Metabolism and immune memory in invertebrates: are they dissociated?Front Immunol. 2024 Jul 11;15:1379471. doi: 10.3389/fimmu.2024.1379471. eCollection 2024. Front Immunol. 2024. PMID: 39055712 Free PMC article. Review.
References
-
- Abdel-Razek AS, Salama HS, White NDG, Morris ON. Effect of Bacillus thuringiensis on feeding and energy use by Plodia interpunctella (Lepidoptera: Pyralidae) and Tribolium castaneum (Coleoptera: Tenebrionidae) The Can Entomol. 1999;131:433–440.
-
- Ali Mohammadie, Kojour M, Han YS, Jo YH. An overview of insect innate immunity. Entomol Res. 2020;50:282–291.
-
- Armitage SA, Peuss R, Kurtz J. Dscam and pancrustacean immune memory-a review of the evidence. Dev Comp Immunol. 2015;48:315–323. - PubMed
-
- Ayres JS, Schneider DS. Tolerance of infections. Annu Rev Immunol. 2012;30:271–294. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
