

The bryophytes Physcomitrium patens and Marchantia polymorpha as model systems for studying evolutionary cell and developmental biology in plants
- PMID: 34459922
- PMCID: PMC8773975
- DOI: 10.1093/plcell/koab218
The bryophytes Physcomitrium patens and Marchantia polymorpha as model systems for studying evolutionary cell and developmental biology in plants
Abstract
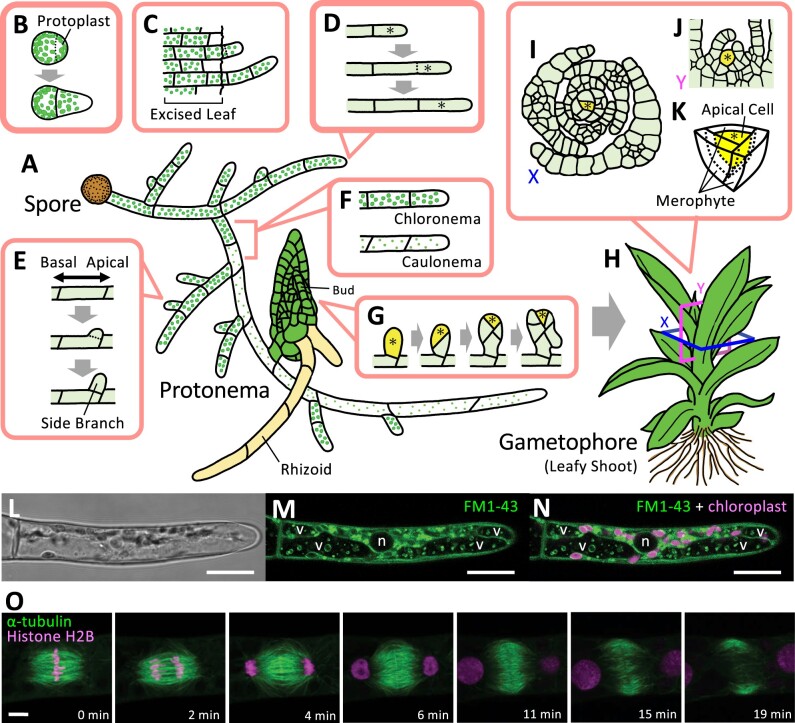
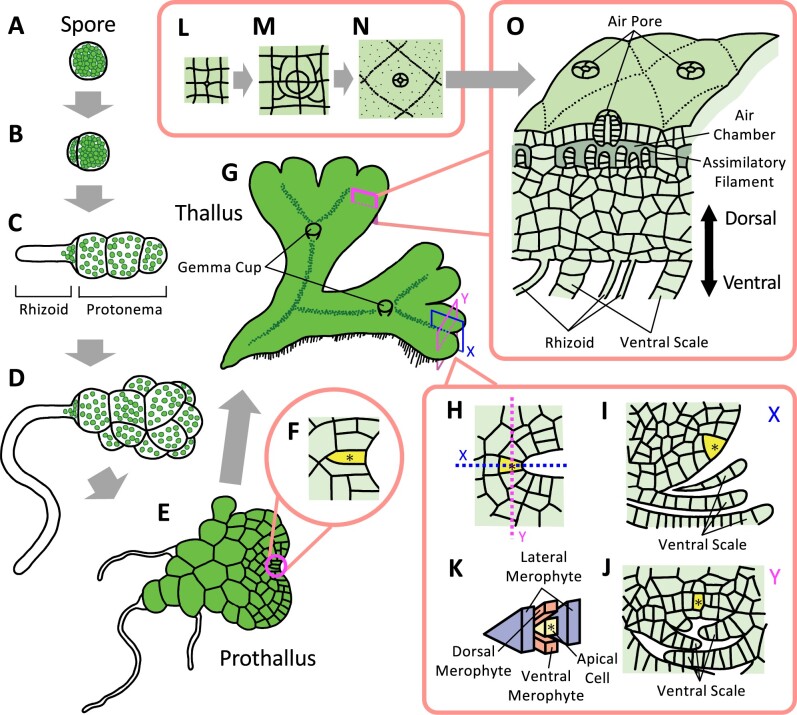
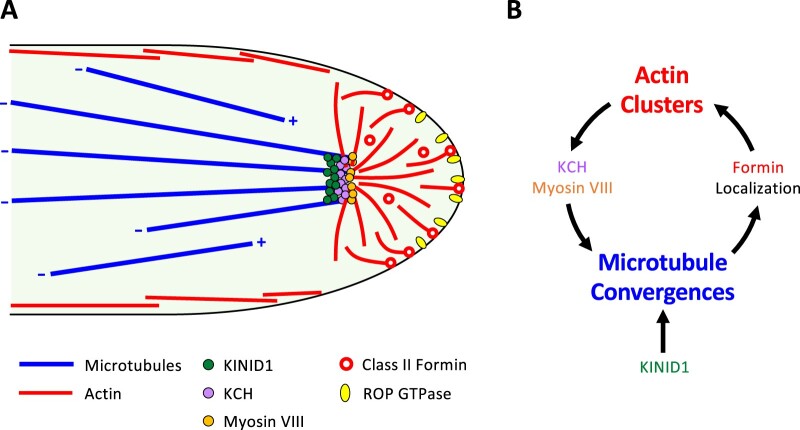
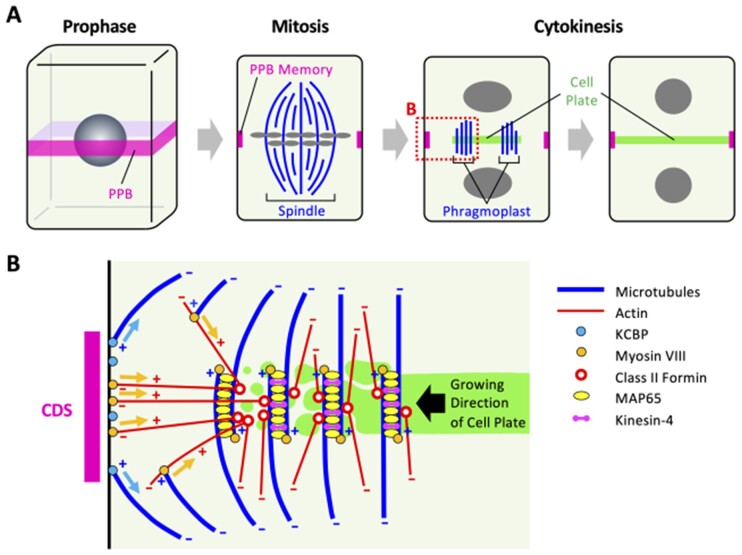
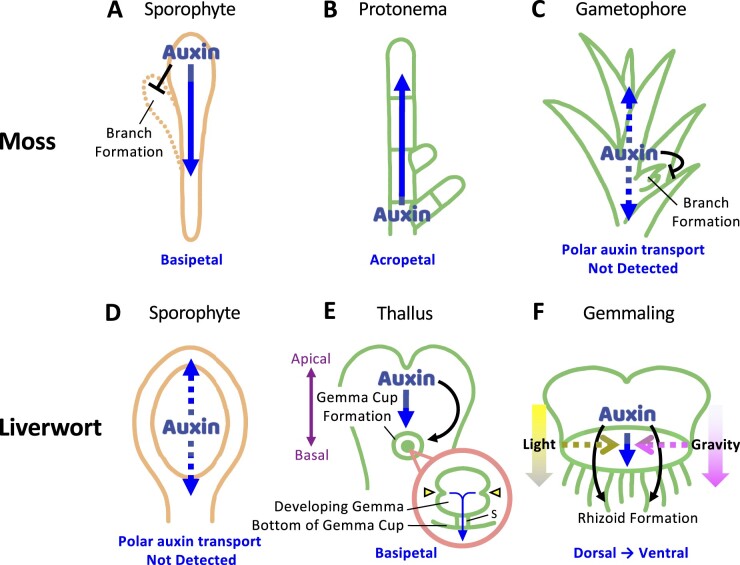
Bryophytes are nonvascular spore-forming plants. Unlike in flowering plants, the gametophyte (haploid) generation of bryophytes dominates the sporophyte (diploid) generation. A comparison of bryophytes with flowering plants allows us to answer some fundamental questions raised in evolutionary cell and developmental biology. The moss Physcomitrium patens was the first bryophyte with a sequenced genome. Many cell and developmental studies have been conducted in this species using gene targeting by homologous recombination. The liverwort Marchantia polymorpha has recently emerged as an excellent model system with low genomic redundancy in most of its regulatory pathways. With the development of molecular genetic tools such as efficient genome editing, both P. patens and M. polymorpha have provided many valuable insights. Here, we review these advances with a special focus on polarity formation at the cell and tissue levels. We examine current knowledge regarding the cellular mechanisms of polarized cell elongation and cell division, including symmetric and asymmetric cell division. We also examine the role of polar auxin transport in mosses and liverworts. Finally, we discuss the future of evolutionary cell and developmental biological studies in plants.
© American Society of Plant Biologists 2021. All rights reserved. For permissions, please email: journals.permissions@oup.com.
Figures
Comment in
-
Back to the roots: A focus on plant cell biology.Plant Cell. 2022 Jan 20;34(1):1-3. doi: 10.1093/plcell/koab278. Plant Cell. 2022. PMID: 34755878 Free PMC article. No abstract available.
References
-
- Allsopp A, Pearman C, Rao AN (1968) The effects of some growth substances and inhibitors on the development of marchantia gemmae. Phytomorphology 18: 84–94
-
- Aoyama T, Hiwatashi Y, Shigyo M, Kofuji R, Kubo M, Ito M, Hasebe M (2012) AP2-type transcription factors determine stem cell identity in the moss Physcomitrella patens. Development 139: 3120–3129 - PubMed
-
- Augustine RC, Vidali L, Kleinman KP, Bezanilla M (2008) Actin depolymerizing factor is essential for viability in plants, and its phosphoregulation is important for tip growth. Plant J 54: 863–875 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
