

Simvastatin modulates estrogen signaling in uterine leiomyoma via regulating receptor palmitoylation, trafficking and degradation
- PMID: 34461224
- PMCID: PMC8455458
- DOI: 10.1016/j.phrs.2021.105856
Simvastatin modulates estrogen signaling in uterine leiomyoma via regulating receptor palmitoylation, trafficking and degradation
Abstract
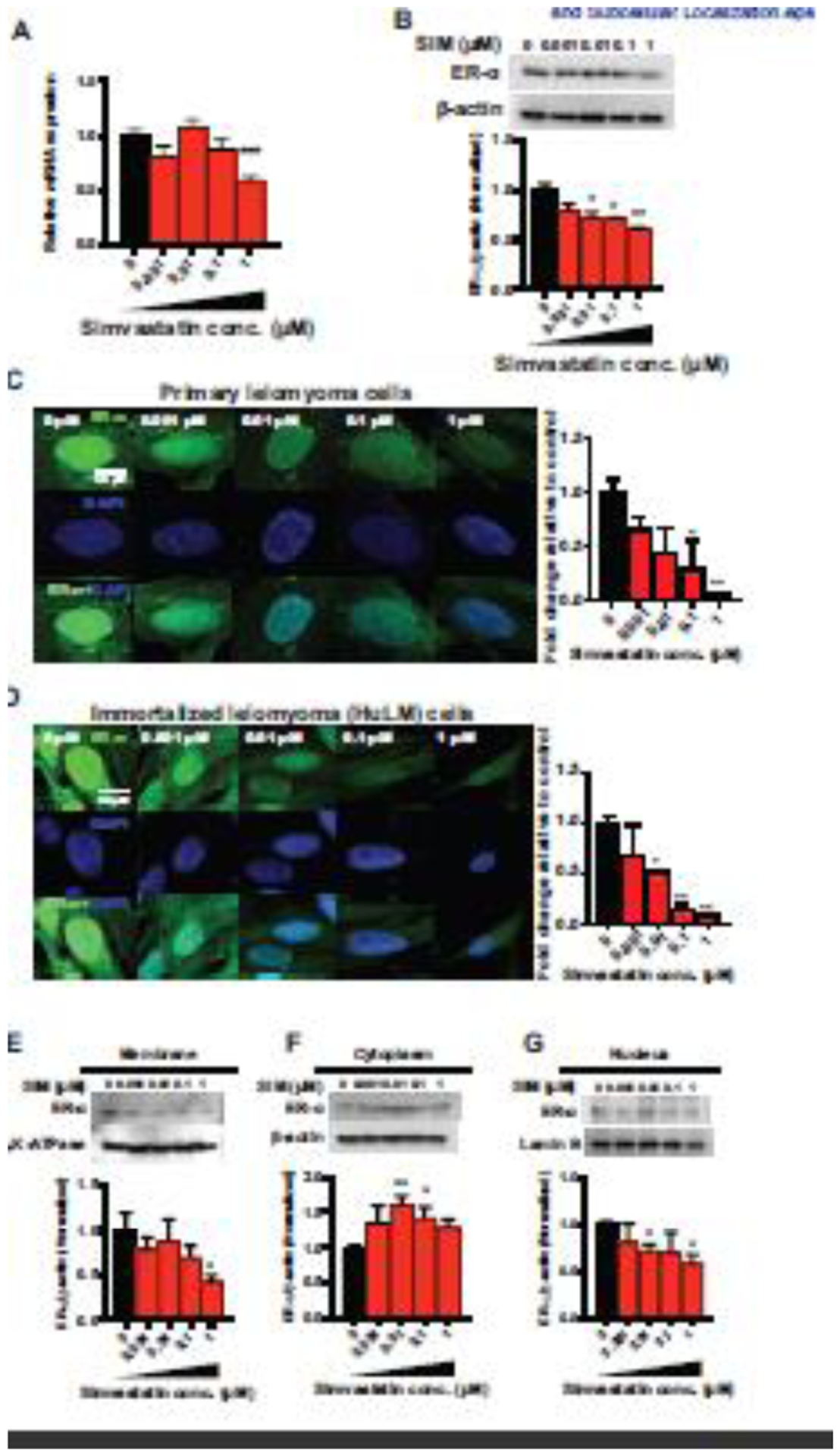
Uterine leiomyomas or fibroids are the most common tumors of the female reproductive tract. Estrogen (E2), a steroid-derived hormone, and its receptors (ERs), particularly ER-α, are important drivers for the development and growth of leiomyomas. We previously demonstrated that simvastatin, a drug used for hyperlipidemia, also possesses anti-leiomyoma properties. The aim of this work is to investigate the impact of simvastatin on ER-α signaling in leiomyoma cells, including its expression, downstream signaling, transcriptional activity, post-translational modification, trafficking and degradation. Primary and immortalized human uterine leiomyoma (HuLM) cells were used for in vitro experiments. Immunodeficient mice xenografted with human leiomyoma tissue explants were used for in vivo studies. Leiomyoma samples were obtained from patients enrolled in an ongoing double-blinded, phase II, randomized controlled trial. Here, we found that simvastatin significantly reduced E2-induced proliferation and PCNA expression. In addition, simvastatin reduced total ER-α expression in leiomyoma cells and altered its subcellular localization by inhibiting its trafficking to the plasma membrane and nucleus. Simvastatin also inhibited E2 downstream signaling, including ERK and AKT pathways, E2/ER transcriptional activity and E2-responsive genes. To explain simvastatin effects on ER-α level and trafficking, we examined its effects on ER-α post-translational processing. We noticed that simvastatin reduced ER-α palmitoylation; a required modification for its stability, trafficking to plasma membrane, and signaling. We also observed an increase in ubiquitin-mediated ER-α degradation. Importantly, we found that the effects of simvastatin on ER-α expression were recapitulated in the xenograft leiomyoma mouse model and human tissues. Thus, our data suggest that simvastatin modulates several E2/ER signaling targets with potential implications in leiomyoma therapy and beyond.
Keywords: Estrogen signaling; Palmitoylation; Receptor degradation; Receptor trafficking; Simvastatin; Uterine leiomyoma.
Copyright © 2021 Elsevier Ltd. All rights reserved.
Conflict of interest statement
Figures






References
-
- Cramer SF, Patel A, The frequency of uterine leiomyomas, Am. J. Clin. Pathol 94(4) (1990) 435–438. - PubMed
-
- Baird DD, Dunson DB, Hill MC, Cousins D, Schectman JM, High cumulative incidence of uterine leiomyoma in black and white women: ultrasound evidence, Am. J. Obstet. Gynecol 188(1) (2003) 100–107. - PubMed
-
- Kasai T, Shozu M, Murakami K, Segawa T, Shinohara K, Nomura K, Inoue M, Increased Expression of Type I 17β-Hydroxysteroid Dehydrogenase Enhances in Situ Production of Estradiol in Uterine Leiomyoma, J. Clin. Endocrinol. Metab 89(11) (2004) 5661–5668. - PubMed
-
- Walker CL, Stewart EA, Uterine fibroids: the elephant in the room, Science 308(5728) (2005) 1589–1592. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
