

AAV integration in human hepatocytes
- PMID: 34461297
- PMCID: PMC8531150
- DOI: 10.1016/j.ymthe.2021.08.031
AAV integration in human hepatocytes
Abstract
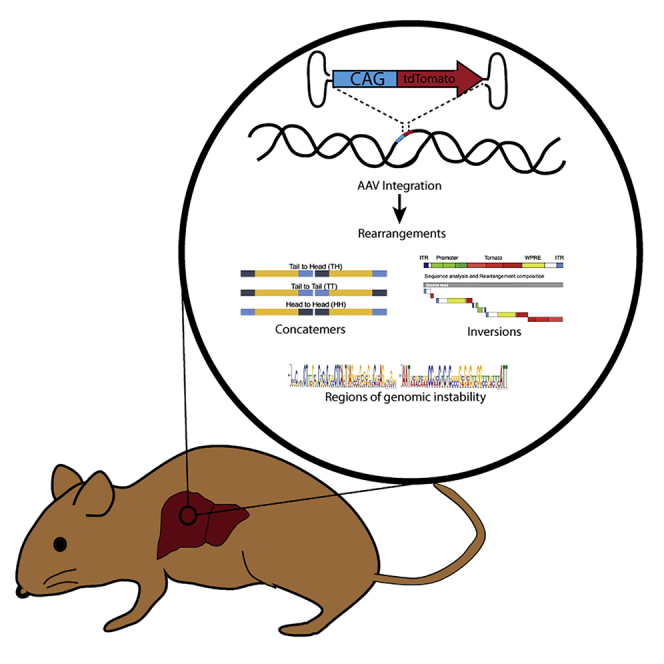
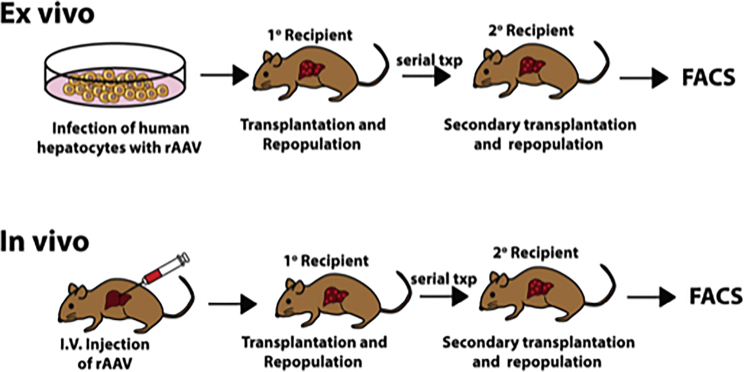
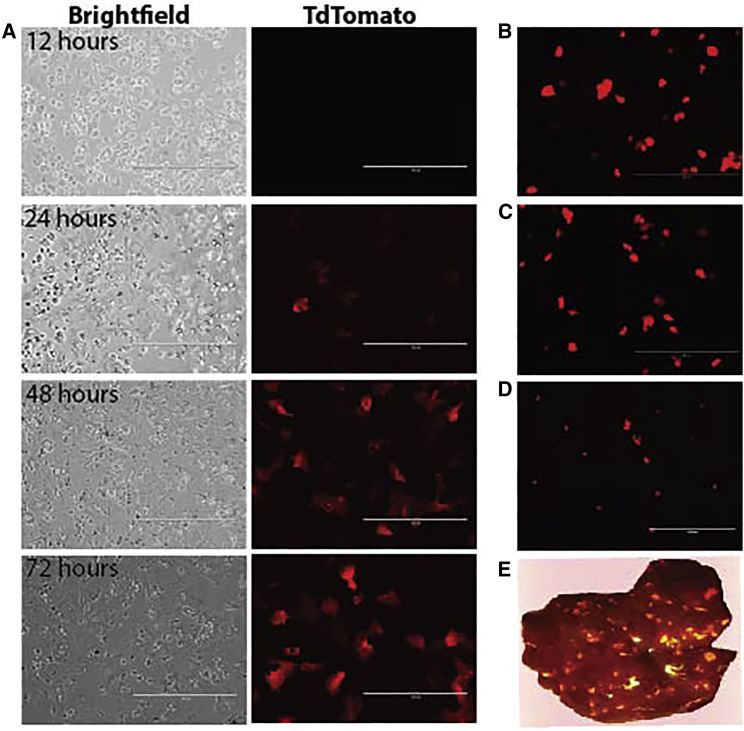
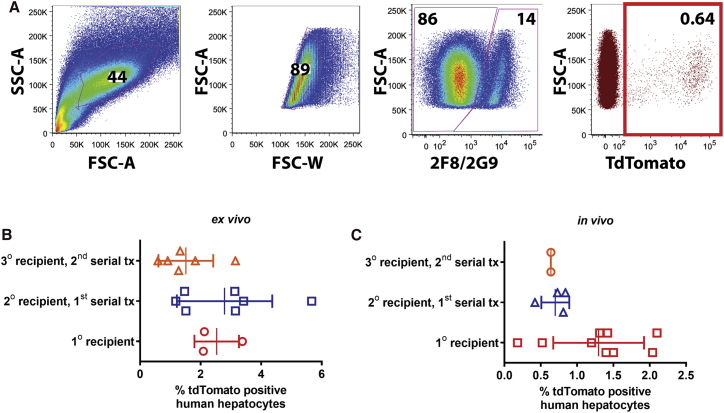
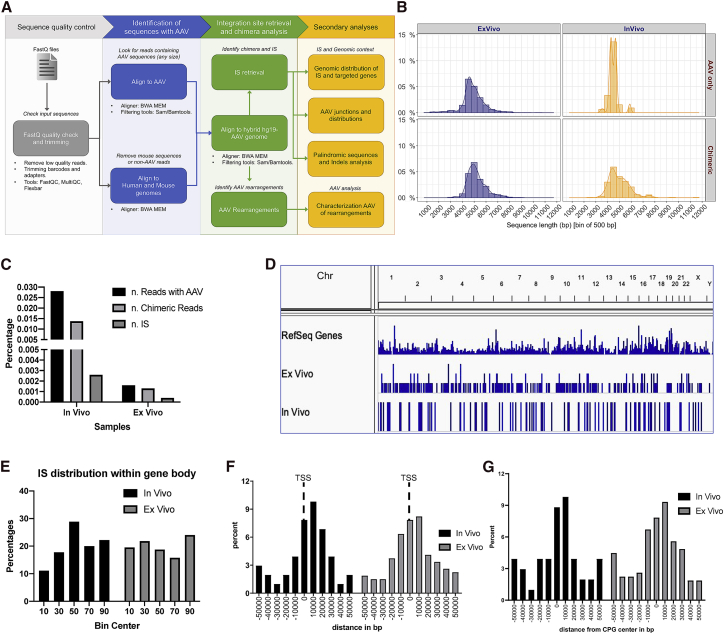
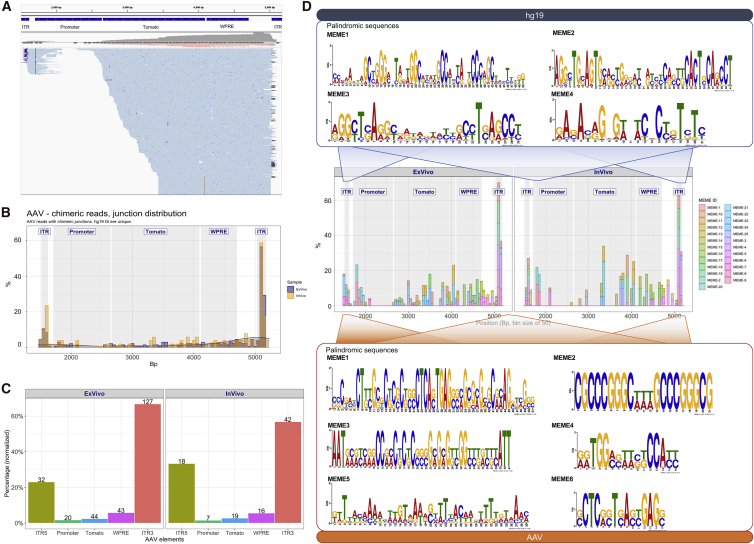
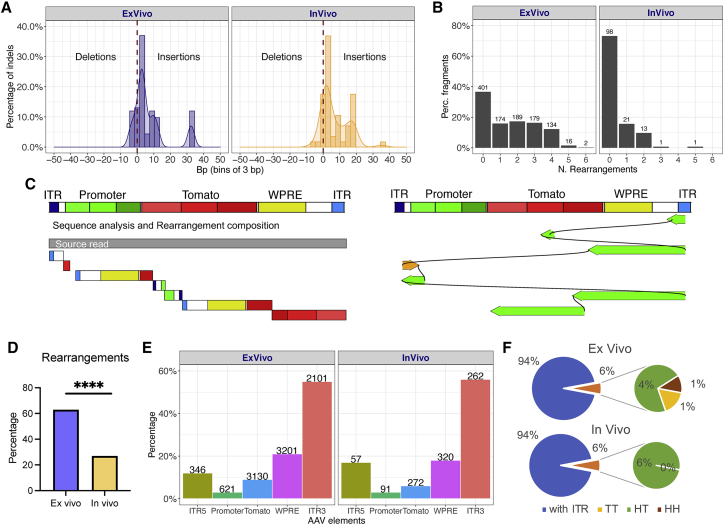
Recombinant adeno-associated viral (rAAV) vectors are considered promising tools for gene therapy directed at the liver. Whereas rAAV is thought to be an episomal vector, its single-stranded DNA genome is prone to intra- and inter-molecular recombination leading to rearrangements and integration into the host cell genome. Here, we ascertained the integration frequency of rAAV in human hepatocytes transduced either ex vivo or in vivo and subsequently expanded in a mouse model of xenogeneic liver regeneration. Chromosomal rAAV integration events and vector integrity were determined using the capture-PacBio sequencing approach, a long-read next-generation sequencing method that has not previously been used for this purpose. Chromosomal integrations were found at a surprisingly high frequency of 1%-3% both in vitro and in vivo. Importantly, most of the inserted rAAV sequences were heavily rearranged and were accompanied by deletions of the host genomic sequence at the integration site.
Keywords: FRGN; capture sequencing; genotoxicity; rAAV; random integration.
Copyright © 2021 The American Society of Gene and Cell Therapy. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests M.G. has a financial interest in Yecuris Corp. (Tigard, Oregon), a company that has commercialized the humanized mouse liver model used in this work. MG also is a consultant for LogicBio Therapeutics. Other authors declare no competing interests.
Figures
References
-
- Anguela X.M., High K.A. Entering the Modern Era of Gene Therapy. Annu. Rev. Med. 2019;70:273–288. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
