

Adult stem cell-derived complete lung organoid models emulate lung disease in COVID-19
- PMID: 34463615
- PMCID: PMC8463074
- DOI: 10.7554/eLife.66417
Adult stem cell-derived complete lung organoid models emulate lung disease in COVID-19
Abstract
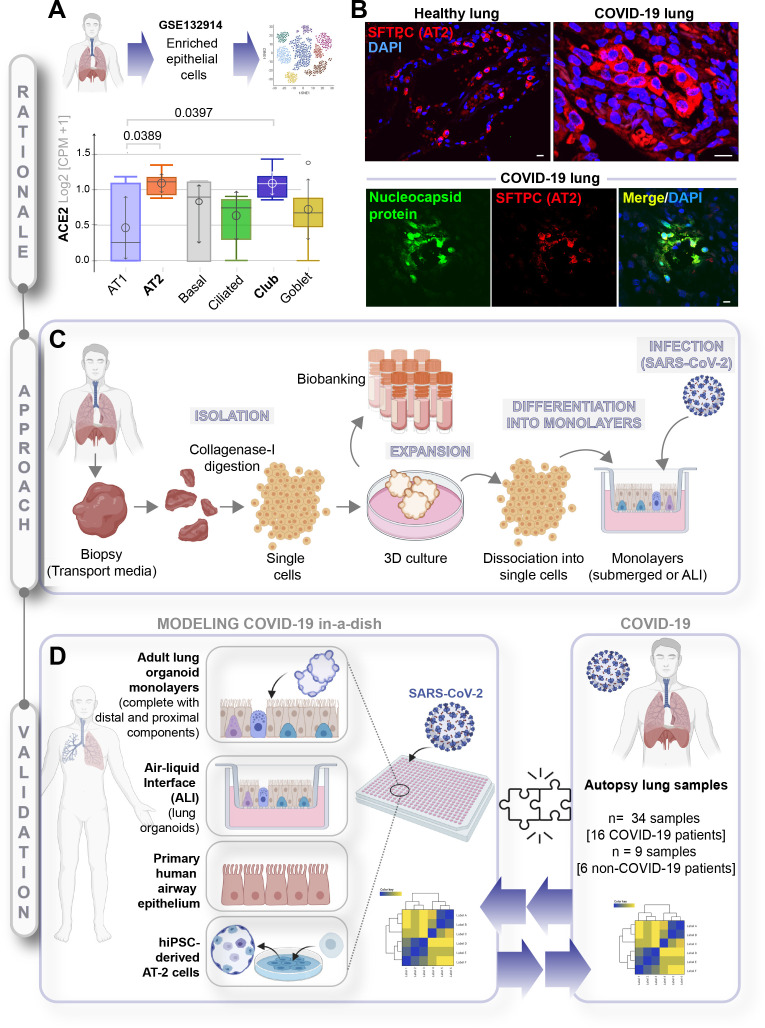
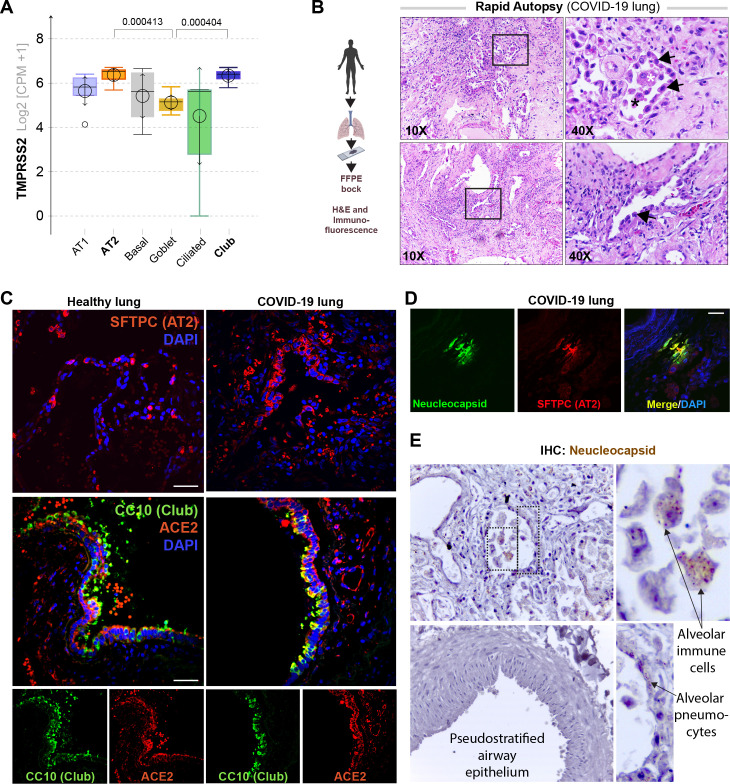
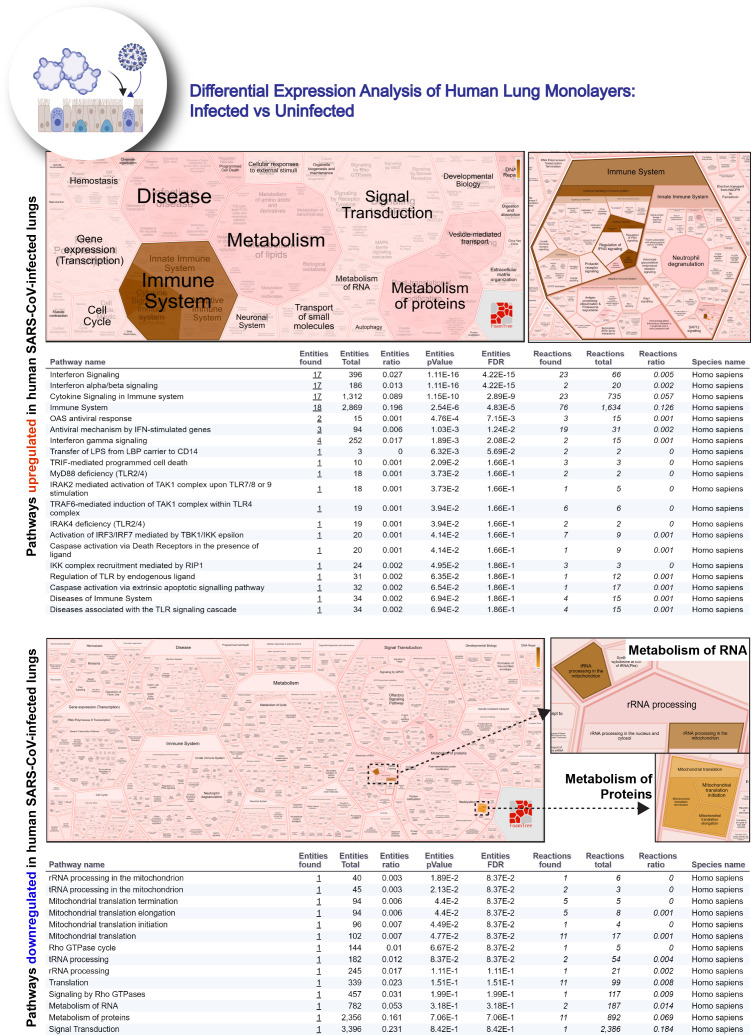
Background: SARS-CoV-2, the virus responsible for COVID-19, causes widespread damage in the lungs in the setting of an overzealous immune response whose origin remains unclear.
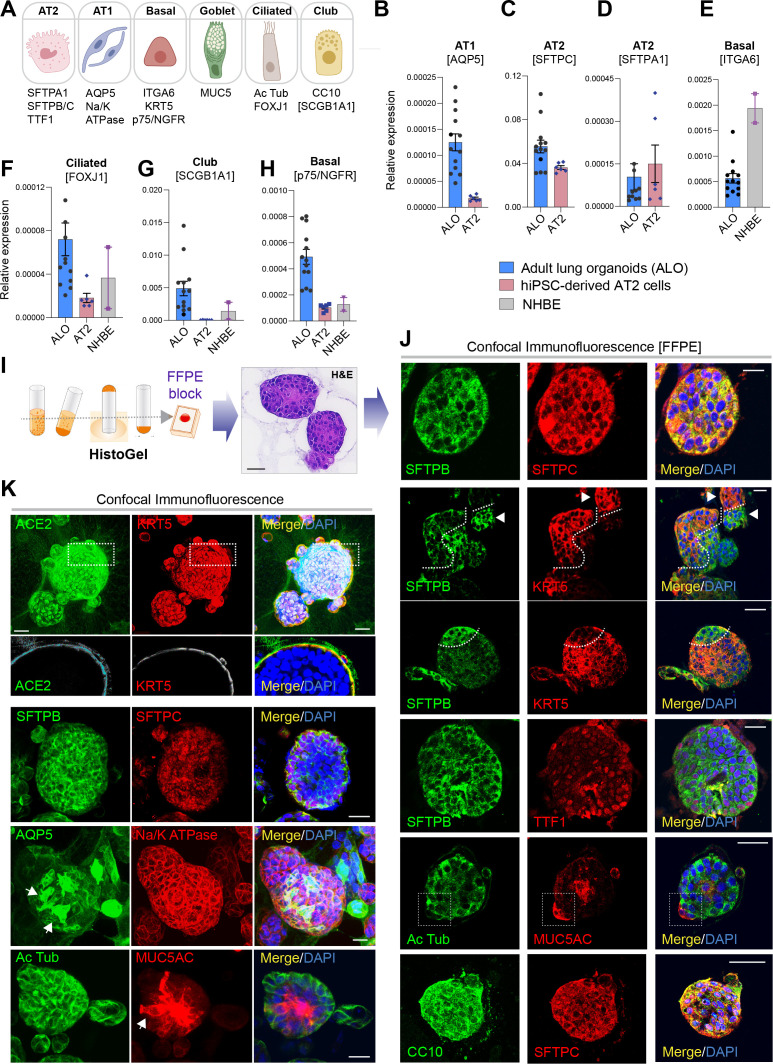
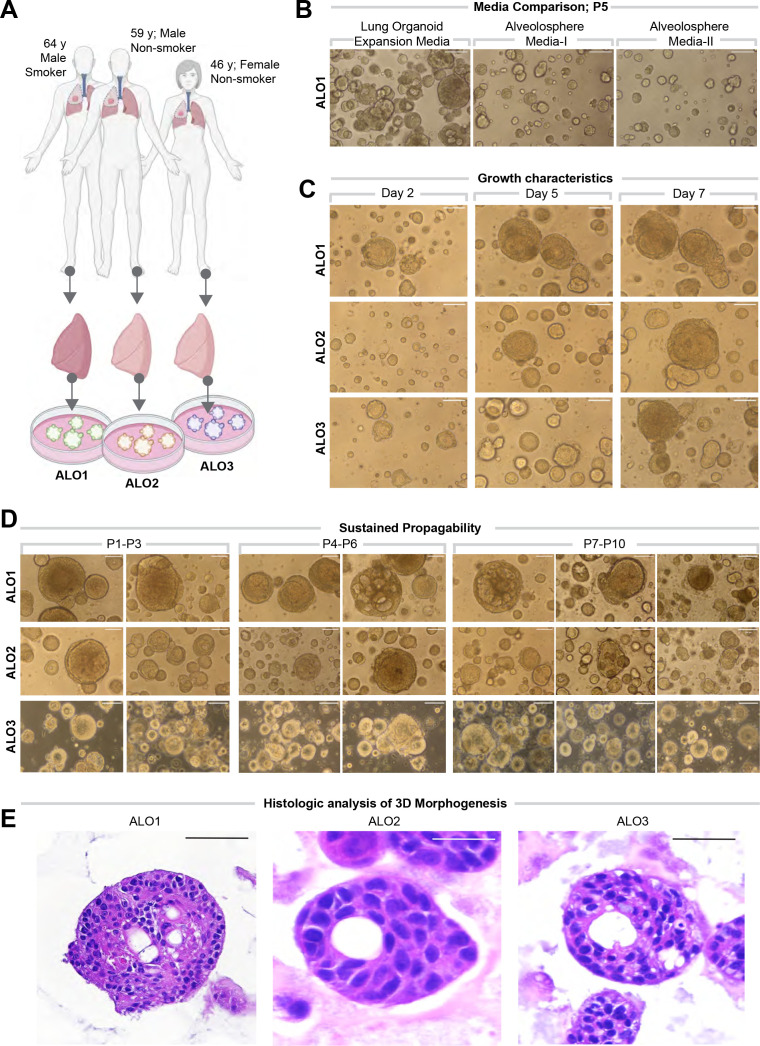
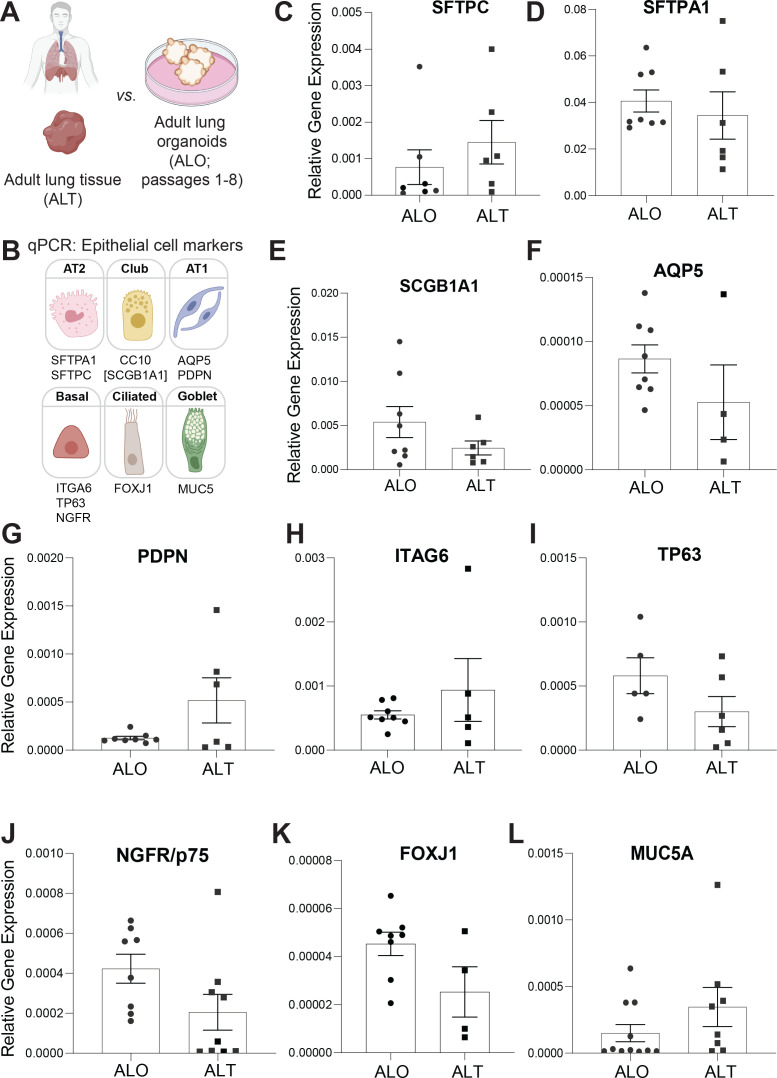
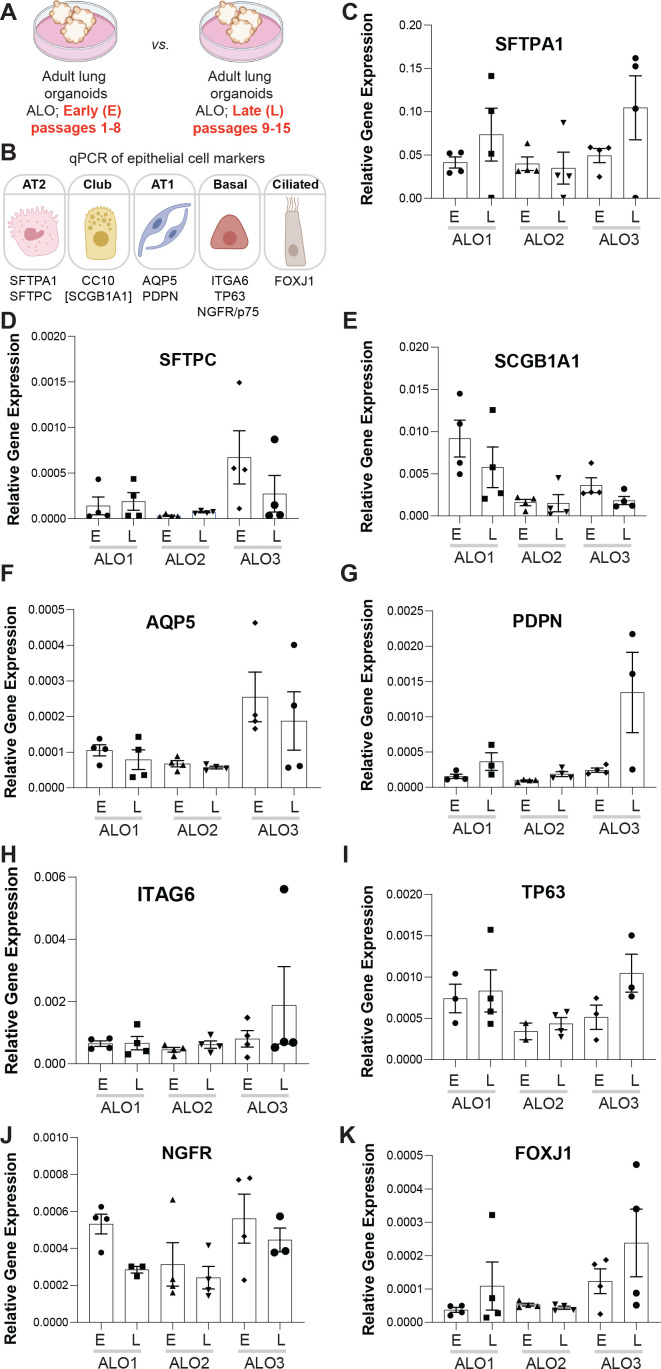
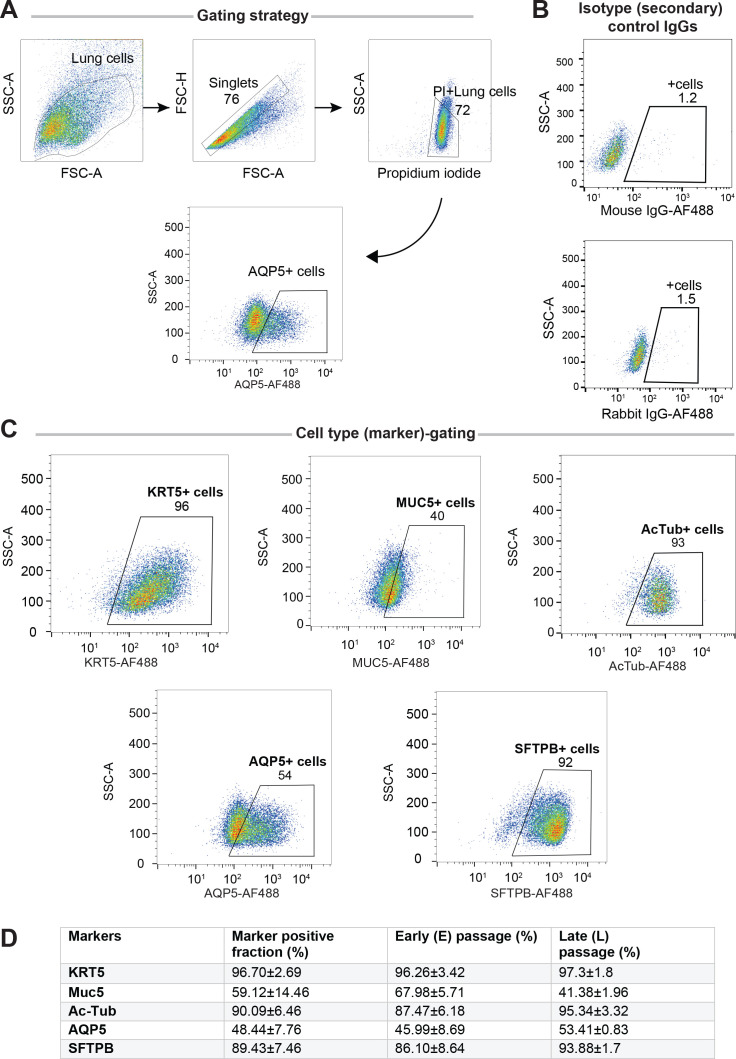
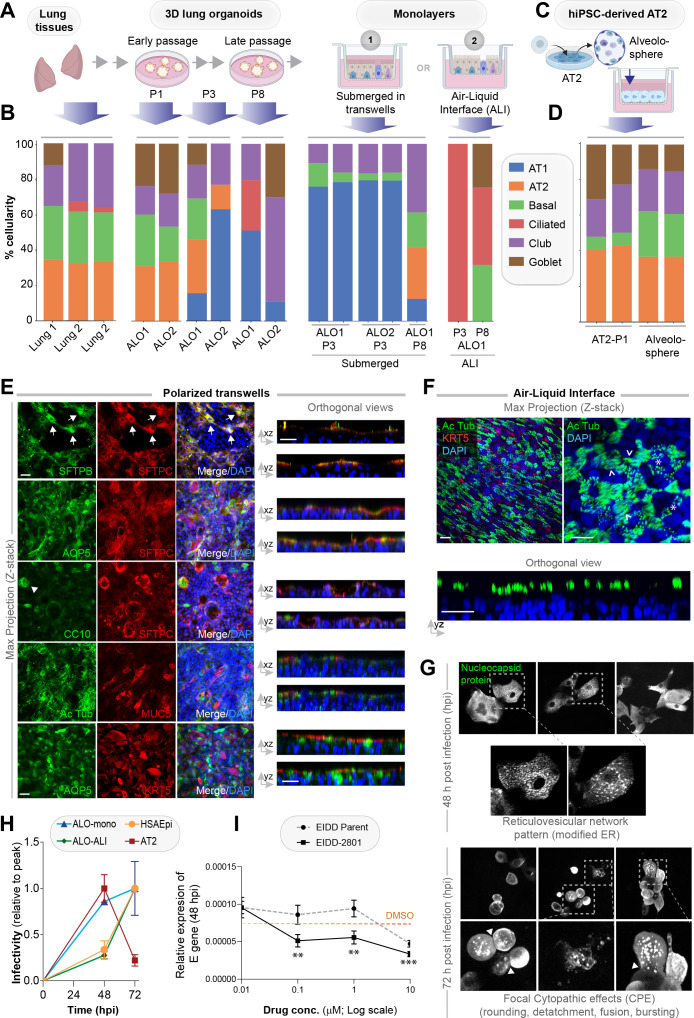
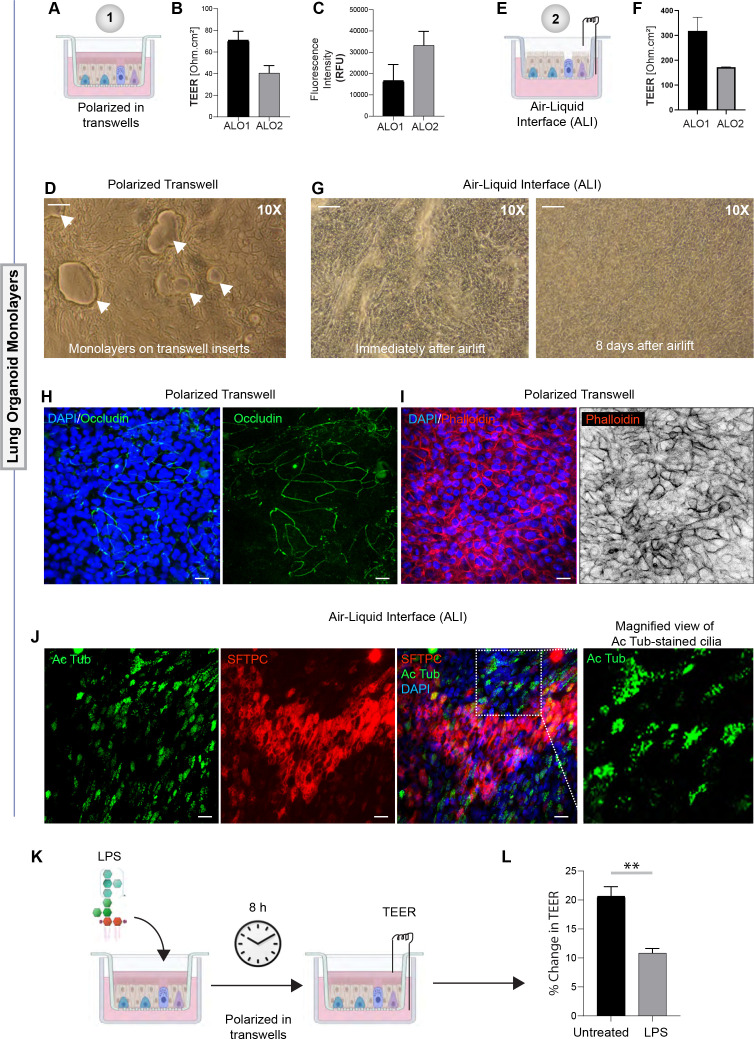
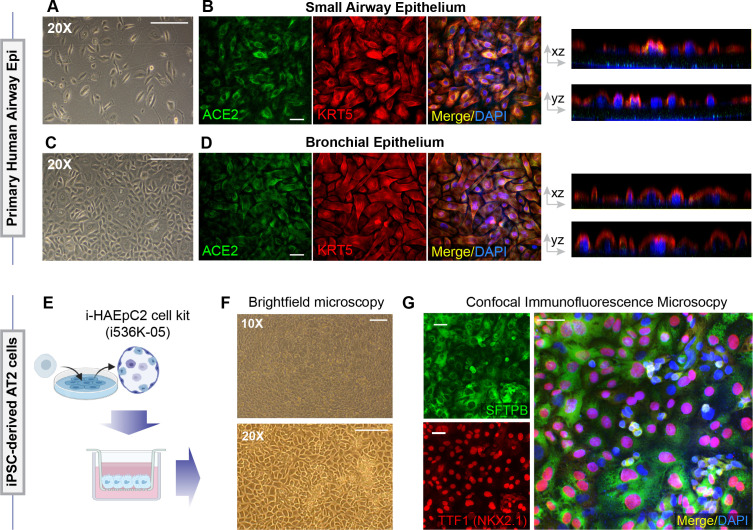
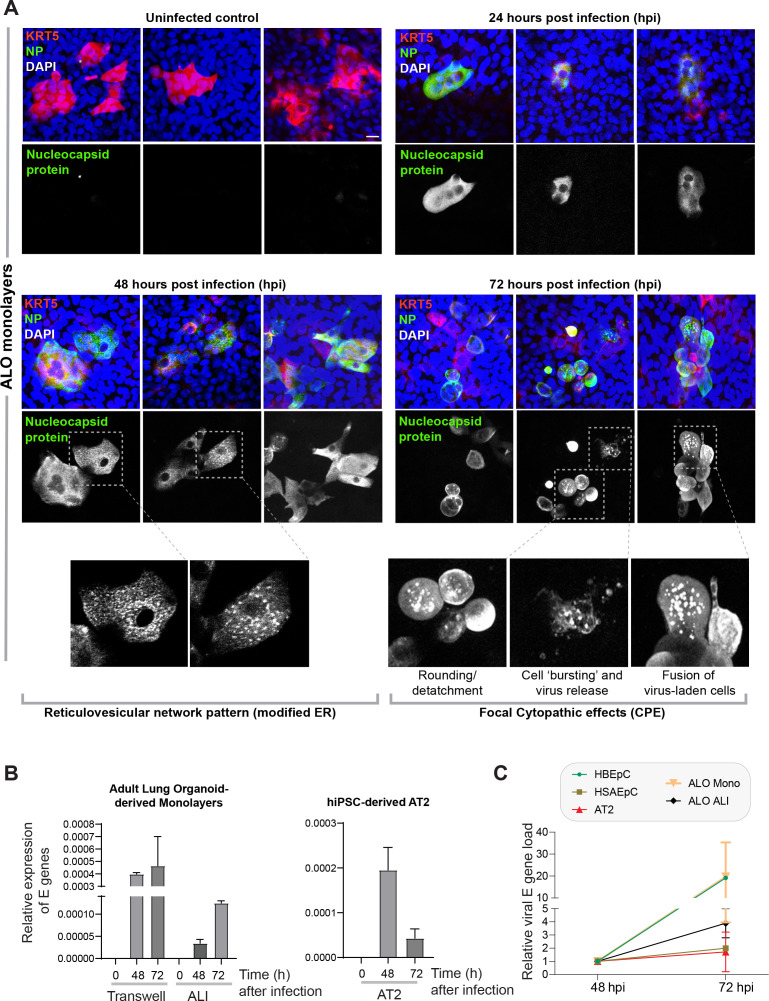
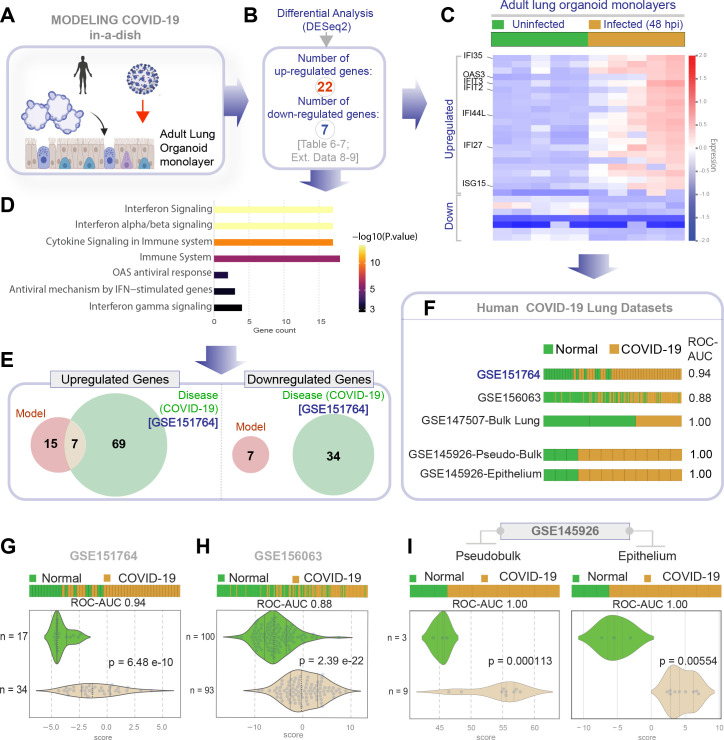
Methods: We present a scalable, propagable, personalized, cost-effective adult stem cell-derived human lung organoid model that is complete with both proximal and distal airway epithelia. Monolayers derived from adult lung organoids (ALOs), primary airway cells, or hiPSC-derived alveolar type II (AT2) pneumocytes were infected with SARS-CoV-2 to create in vitro lung models of COVID-19.
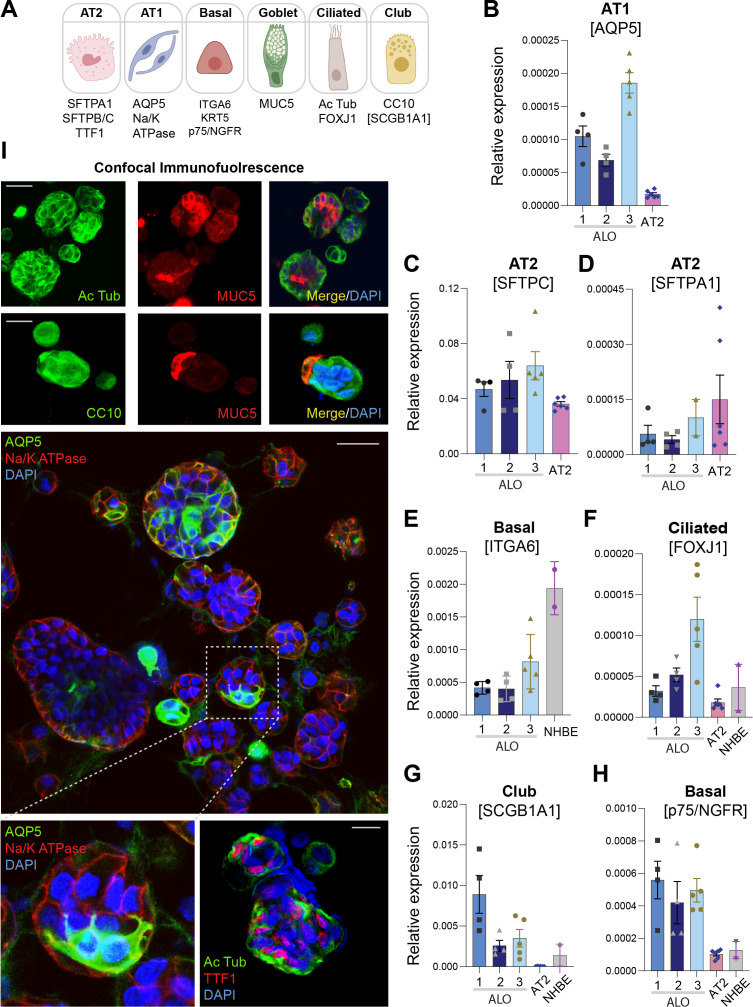
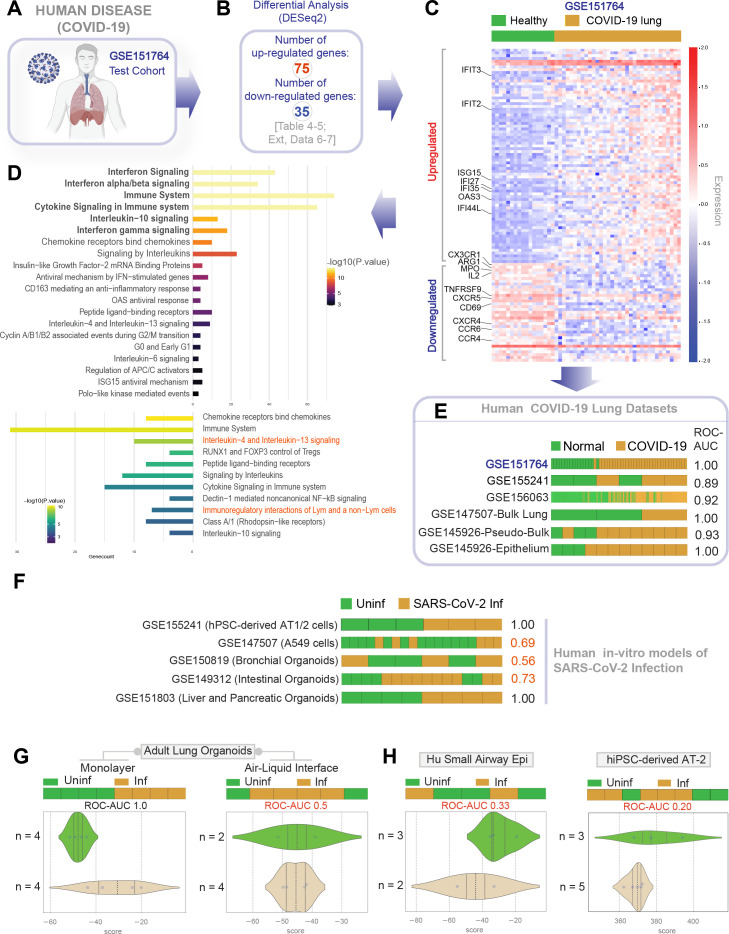
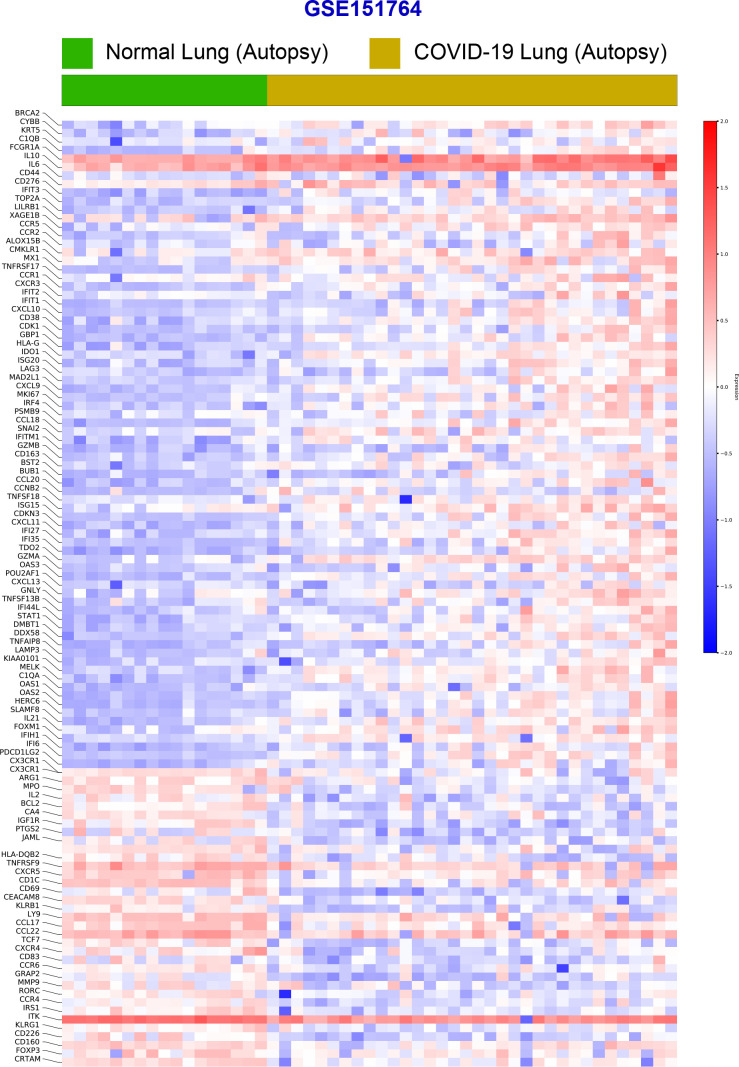
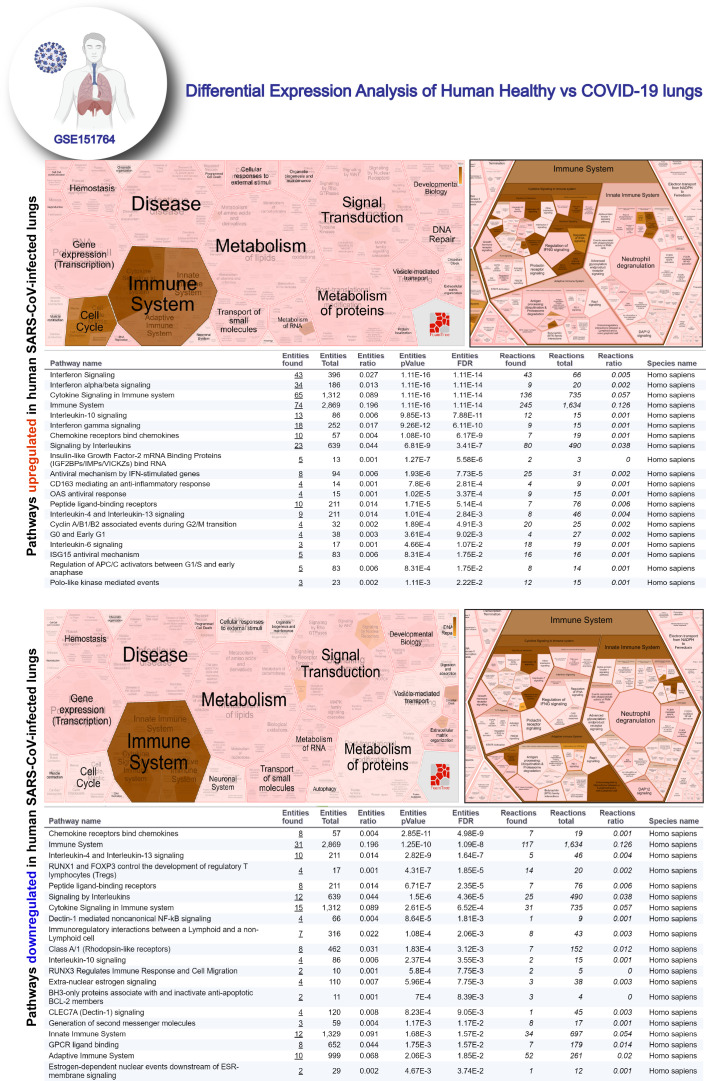
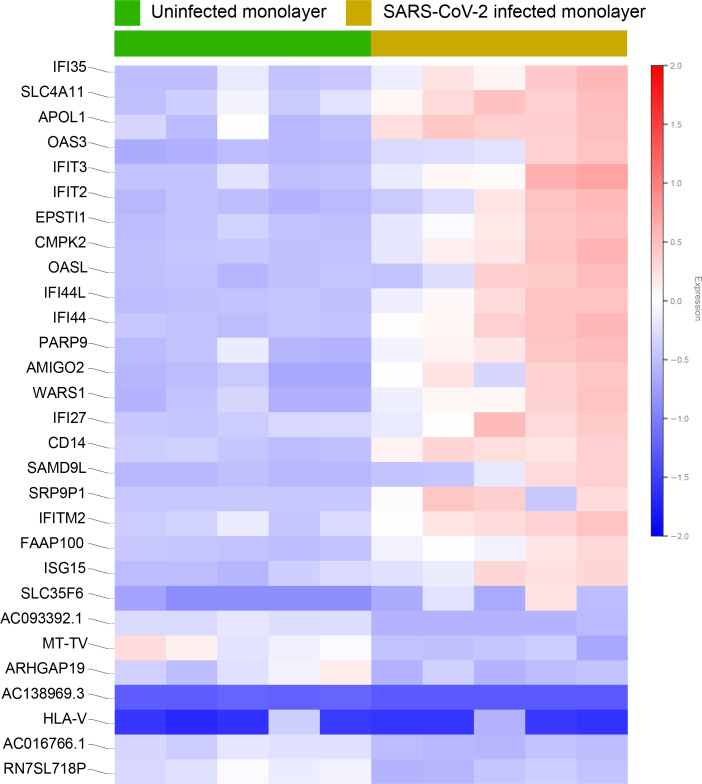
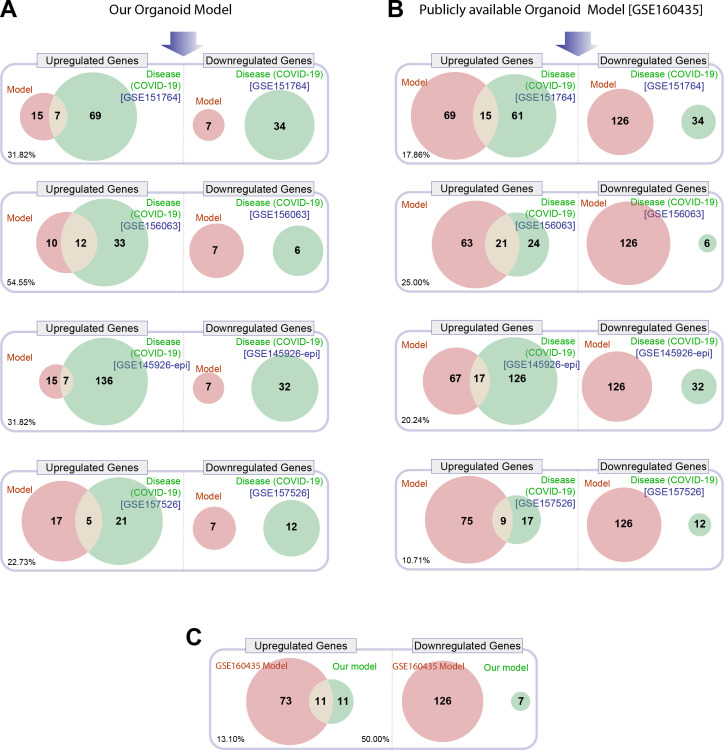
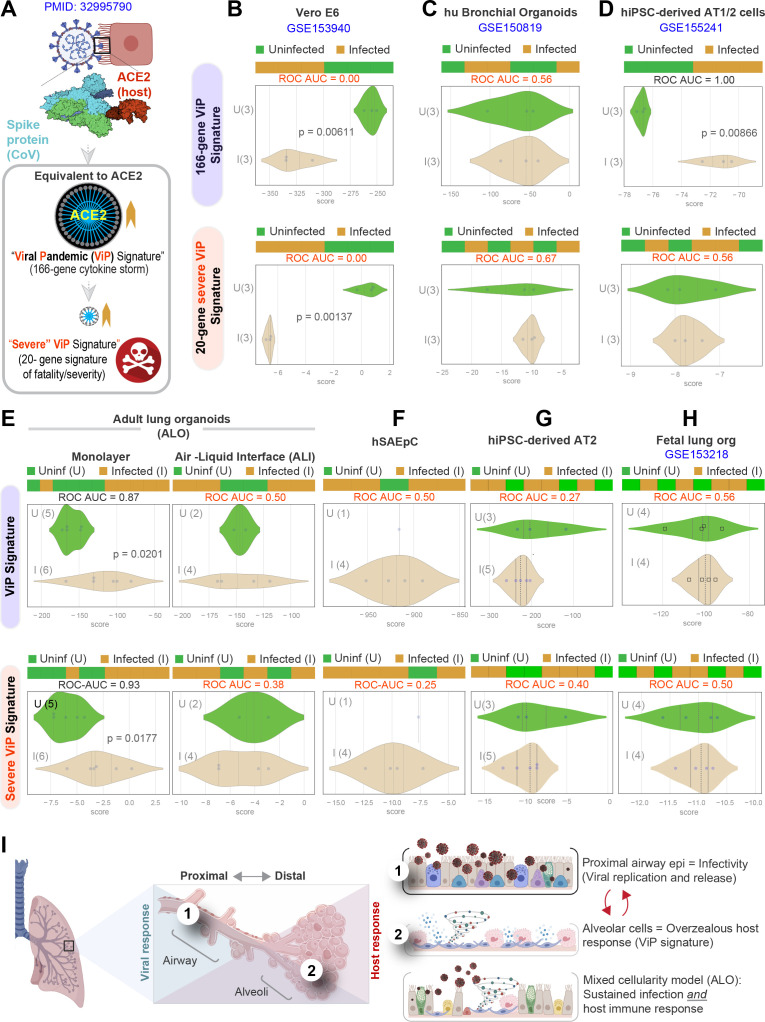
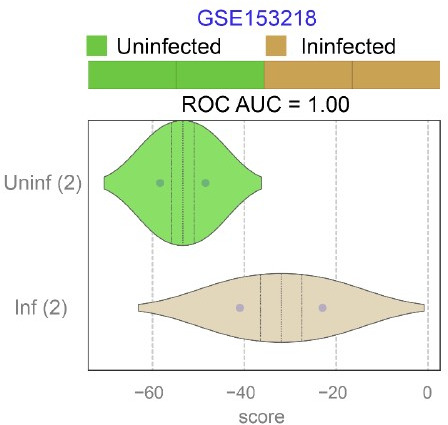
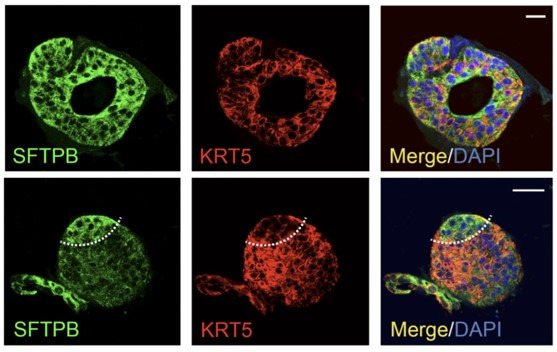
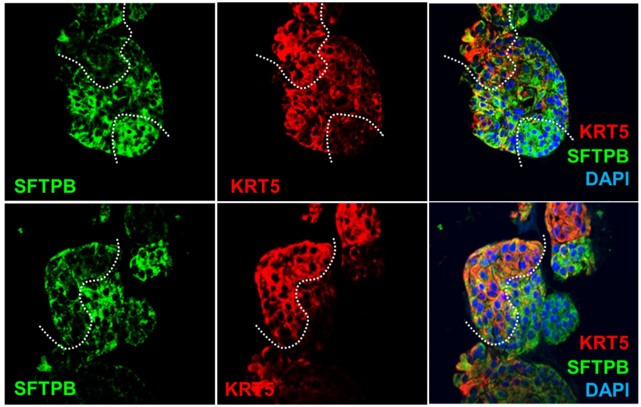
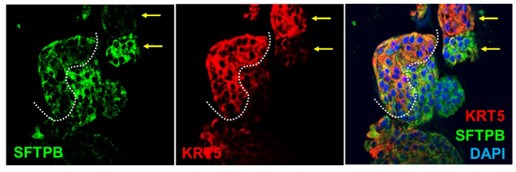
Results: Infected ALO monolayers best recapitulated the transcriptomic signatures in diverse cohorts of COVID-19 patient-derived respiratory samples. The airway (proximal) cells were critical for sustained viral infection, whereas distal alveolar differentiation (AT2→AT1) was critical for mounting the overzealous host immune response in fatal disease; ALO monolayers with well-mixed proximodistal airway components recapitulated both.
Conclusions: Findings validate a human lung model of COVID-19, which can be immediately utilized to investigate COVID-19 pathogenesis and vet new therapies and vaccines.
Funding: This work was supported by the National Institutes for Health (NIH) grants 1R01DK107585-01A1, 3R01DK107585-05S1 (to SD); R01-AI141630, CA100768 and CA160911 (to PG) and R01-AI 155696 (to PG, DS and SD); R00-CA151673 and R01-GM138385 (to DS), R01- HL32225 (to PT), UCOP-R00RG2642 (to SD and PG), UCOP-R01RG3780 (to P.G. and D.S) and a pilot award from the Sanford Stem Cell Clinical Center at UC San Diego Health (P.G, S.D, D.S). GDK was supported through The American Association of Immunologists Intersect Fellowship Program for Computational Scientists and Immunologists. L.C.A's salary was supported in part by the VA San Diego Healthcare System. This manuscript includes data generated at the UC San Diego Institute of Genomic Medicine (IGC) using an Illumina NovaSeq 6000 that was purchased with funding from a National Institutes of Health SIG grant (#S10 OD026929).
Keywords: AT2 differentiation; SARS-CoV2; computational; disease modeling; human; immune response; lung organoid; medicine; regenerative medicine; stem cells; viruses.
© 2021, Tindle et al.
Conflict of interest statement
CT, AF, ST, SI, NB, GK, AC, VC, MH, HR, JD, LC, AT, GL, PT, RC, TR, DS, PG, SD None, MF none
Figures
Update of
-
Adult Stem Cell-derived Complete Lung Organoid Models Emulate Lung Disease in COVID-19.bioRxiv [Preprint]. 2021 May 5:2020.10.17.344002. doi: 10.1101/2020.10.17.344002. bioRxiv. 2021. Update in: Elife. 2021 Aug 13;10:e66417. doi: 10.7554/eLife.66417. PMID: 33106807 Free PMC article. Updated. Preprint.
References
-
- Abdelwahab EMM, Rapp J, Feller D, Csongei V, Pal S, Bartis D, Thickett DR, Pongracz JE. WNT signaling regulates trans-differentiation of stem cell like type 2 alveolar epithelial cells to type 1 epithelial cells. Respiratory Research. 2019;20:204. doi: 10.1186/s12931-019-1176-x. - DOI - PMC - PubMed
-
- Barrett T, Wilhite SE, Ledoux P, Evangelista C, Kim IF, Tomashevsky M, Marshall KA, Phillippy KH, Sherman PM, Holko M, Yefanov A, Lee H, Zhang N, Robertson CL, Serova N, Davis S, Soboleva A. NCBI GEO: Archive for functional Genomics data sets--update. Nucleic Acids Research. 2013;41:D991–D995. doi: 10.1093/nar/gks1193. - DOI - PMC - PubMed
-
- Bonser LR, Koh KD, Johansson K, Choksi SP, Cheng D, Liu L, Sun DI, Zlock LT, Eckalbar WL, Finkbeiner WE, Erle DJ. Flow-cytometric analysis and purification of airway epithelial-cell subsets. American Journal of Respiratory Cell and Molecular Biology. 2021;64:308–317. doi: 10.1165/rcmb.2020-0149MA. - DOI - PMC - PubMed
Publication types
MeSH terms
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
