

Osteoblastic glucocorticoid signaling exacerbates high-fat-diet- induced bone loss and obesity
- PMID: 34465731
- PMCID: PMC8408138
- DOI: 10.1038/s41413-021-00159-9
Osteoblastic glucocorticoid signaling exacerbates high-fat-diet- induced bone loss and obesity
Abstract
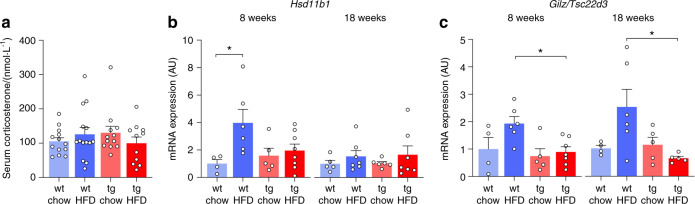
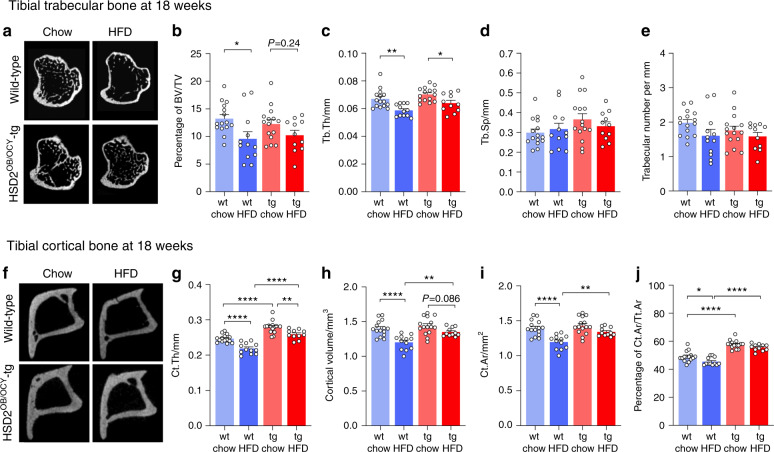
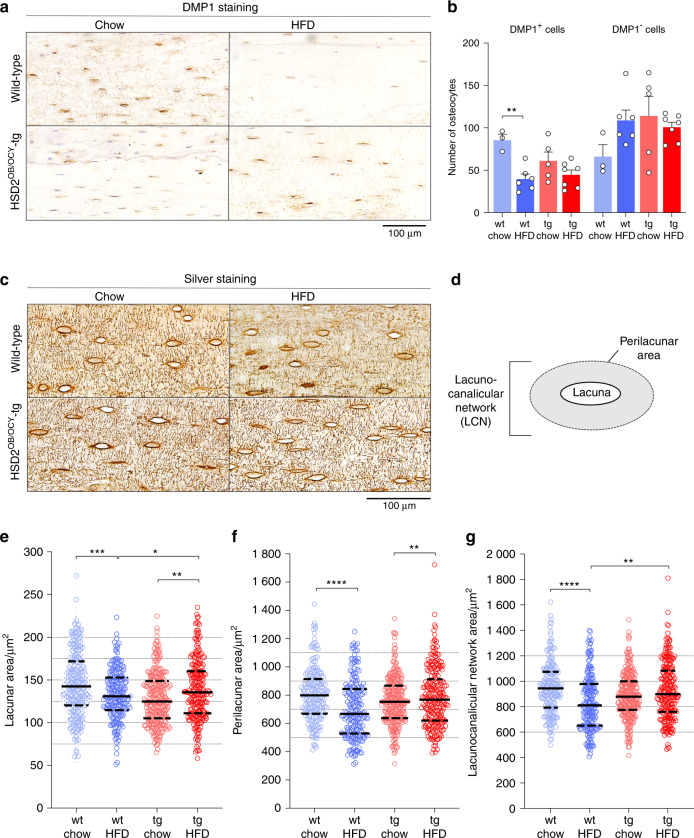
Chronic high-fat diet (HFD) consumption not only promotes obesity and insulin resistance, but also causes bone loss through mechanisms that are not well understood. Here, we fed wild-type CD-1 mice either chow or a HFD (43% of energy from fat) for 18 weeks; HFD-fed mice exhibited decreased trabecular volume (-28%) and cortical thickness (-14%) compared to chow-fed mice. In HFD-fed mice, bone loss was due to reduced bone formation and mineral apposition, without obvious effects on bone resorption. HFD feeding also increased skeletal expression of sclerostin and caused deterioration of the osteocyte lacunocanalicular network (LCN). In mice fed HFD, skeletal glucocorticoid signaling was activated relative to chow-fed mice, independent of serum corticosterone concentrations. We therefore examined whether skeletal glucocorticoid signaling was necessary for HFD-induced bone loss, using transgenic mice lacking glucocorticoid signaling in osteoblasts and osteocytes (HSD2OB/OCY-tg mice). In HSD2OB/OCY-tg mice, bone formation and mineral apposition rates were not suppressed by HFD, and bone loss was significantly attenuated. Interestingly, in HSD2OB/OCY-tg mice fed HFD, both Wnt signaling (less sclerostin induction, increased β-catenin expression) and glucose uptake were significantly increased, relative to diet- and genotype-matched controls. The osteocyte LCN remained intact in HFD-fed HSD2OB/OCY-tg mice. When fed a HFD, HSD2OB/OCY-tg mice also increased their energy expenditure and were protected against obesity, insulin resistance, and dyslipidemia. Therefore, glucocorticoid signaling in osteoblasts and osteocytes contributes to the suppression of bone formation in HFD-fed mice. Skeletal glucocorticoid signaling is also an important determinant of glucose uptake in bone, which influences the whole-body metabolic response to HFD.
© 2021. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures




References
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
