

Skeletal Muscle Is an Early Site of Zika Virus Replication and Injury, Which Impairs Myogenesis
- PMID: 34468171
- PMCID: PMC8549499
- DOI: 10.1128/JVI.00904-21
Skeletal Muscle Is an Early Site of Zika Virus Replication and Injury, Which Impairs Myogenesis
Abstract
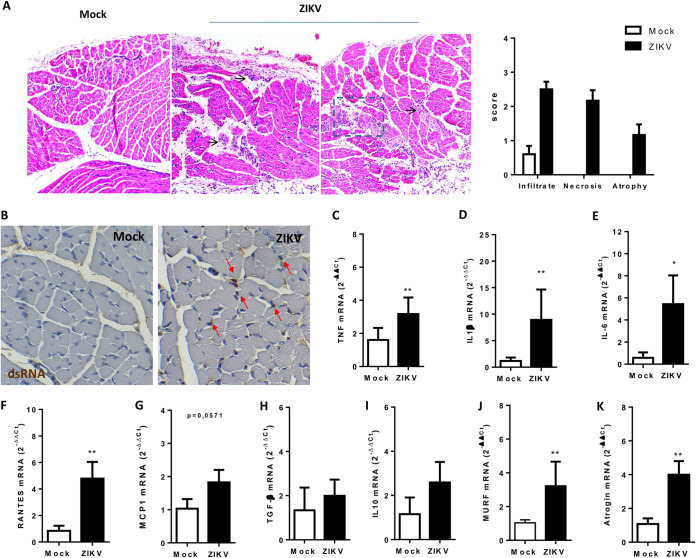
Zika virus (ZIKV) infection became a worldwide concern due to its correlation with the development of microcephaly and other neurological disorders. ZIKV neurotropism is well characterized, but the role of peripheral viral amplification to brain infection remains unknown. Here, we found that ZIKV replicates in human primary skeletal muscle myoblasts, impairing its differentiation into myotubes but not interfering with the integrity of the already-formed muscle fibers. Using mouse models, we showed ZIKV tropism to muscle tissue either during embryogenesis after maternal transmission or when infection occurred after birth. Interestingly, ZIKV replication in the mouse skeletal muscle started immediately after ZIKV inoculation, preceding viral RNA detection in the brain and causing no disruption to the integrity of the blood brain barrier, and remained active for more than 2 weeks, whereas replication in the spleen and liver were not sustained over time. In addition, ZIKV infection of the skeletal muscle induces necrotic lesions, inflammation, and fiber atrophy. We also found a reduction in the expression of regulatory myogenic factors that are essential for muscle repair after injury. Taken together, our results indicate that the skeletal muscle is an early site of viral amplification and lesion that may result in late consequences in muscle development after ZIKV infection. IMPORTANCE Zika Virus (ZIKV) neurotropism and its deleterious effects on central nervous system have been well characterized. However, investigations of the initial replication sites for the establishment of infection and viral spread to neural tissues remain underexplored. A complete description of the range of ZIKV-induced lesions and others factors that can influence the severity of the disease is necessary to prevent ZIKV's deleterious effects. ZIKV has been shown to access the central nervous system without significantly affecting blood-brain barrier permeability. Here, we demonstrated that skeletal muscle is an earlier site of ZIKV replication, contributing to the increase of peripheral ZIKV load. ZIKV replication in muscle promotes necrotic lesions and inflammation and also impairs myogenesis. Overall, our findings showed that skeletal muscle is involved in pathogenesis and opens new fields in the investigation of the long-term consequences of early infection.
Keywords: Zika virus replication; muscle inflammation; myogenesis; pathogenesis; skeletal muscle; viral dissemination.
Figures





References
-
- Krauer F, Riesen M, Reveiz L, Oladapo OT, Martinez-Vega R, Porgo TV, Haefliger A, Broutet NJ, Low N, Group W, WHO Zika Causality Working Group. 2017. Zika virus infection as a cause of congenital brain abnormalities and Guillain-Barre syndrome: systematic review. PLoS Med 14:e1002203. 10.1371/journal.pmed.1002203. - DOI - PMC - PubMed
Publication types
MeSH terms
Grants and funding
- LCF/PR/HR17/52150011/"la Caixa" Foundation ("la Caixa")
- 203.225/2017/Fundação Carlos Chagas Filho de Amparo à Pesquisa do Estado do Rio de Janeiro (FAPERJ)
- 201.316/2016/Fundação Carlos Chagas Filho de Amparo à Pesquisa do Estado do Rio de Janeiro (FAPERJ)
- 309028/2017-5/MCTI | Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq)
- 436933/2018-7/MCTI | Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq)
LinkOut - more resources
Full Text Sources
Medical
