

IL-33 causes thermogenic failure in aging by expanding dysfunctional adipose ILC2
- PMID: 34473956
- PMCID: PMC9067336
- DOI: 10.1016/j.cmet.2021.08.004
IL-33 causes thermogenic failure in aging by expanding dysfunctional adipose ILC2
Abstract
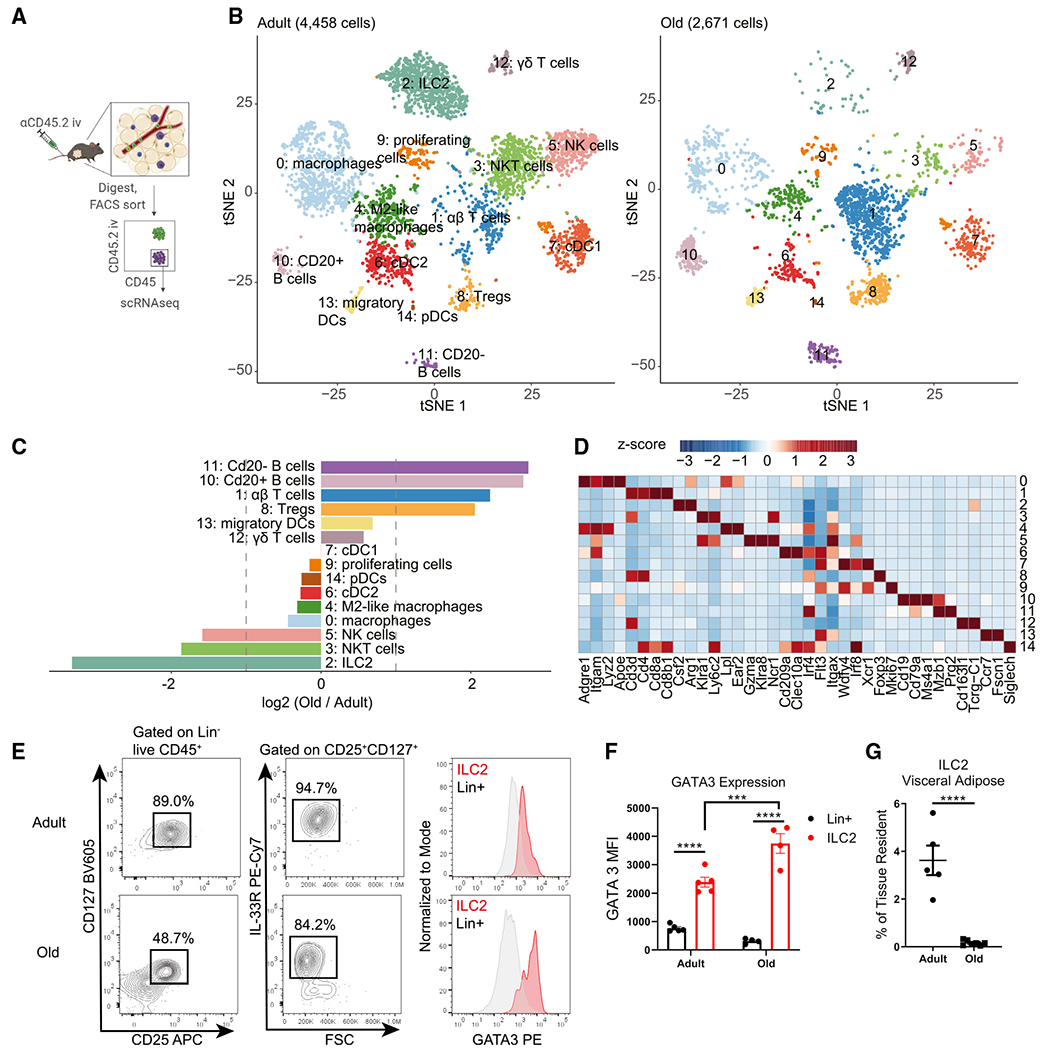
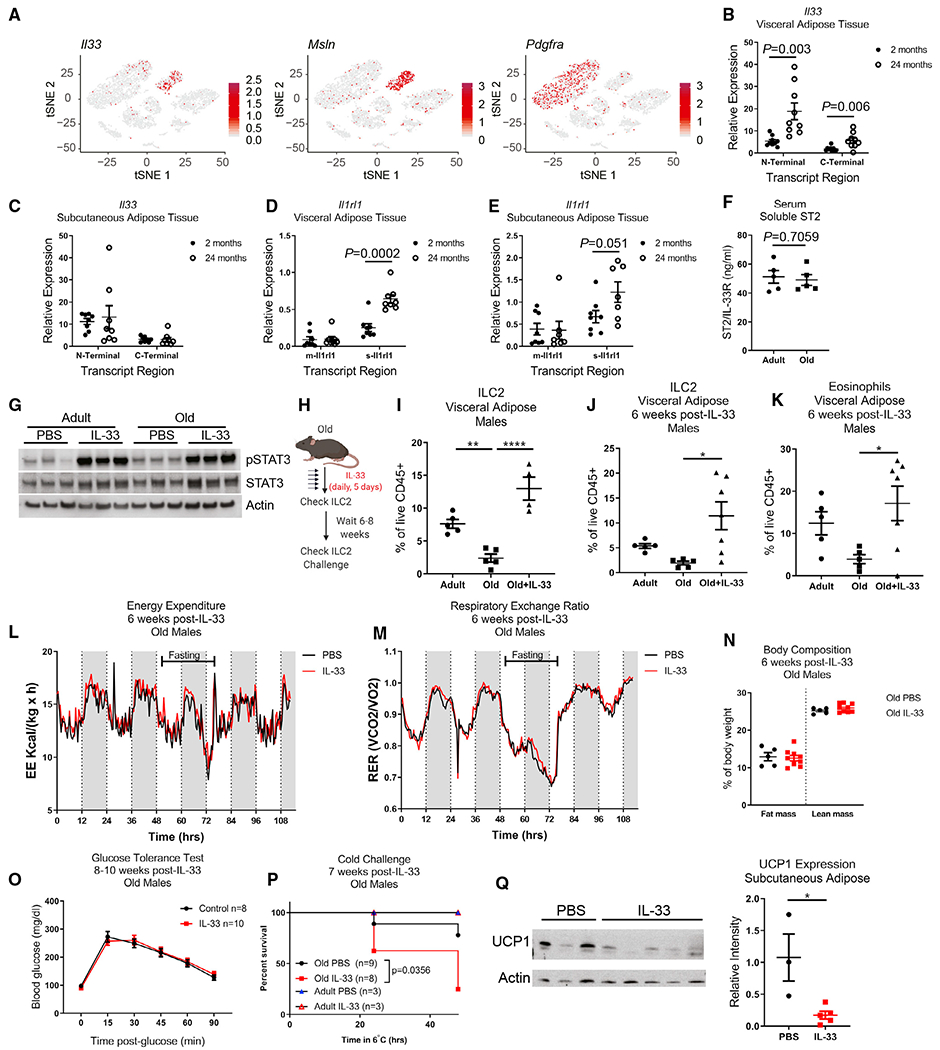
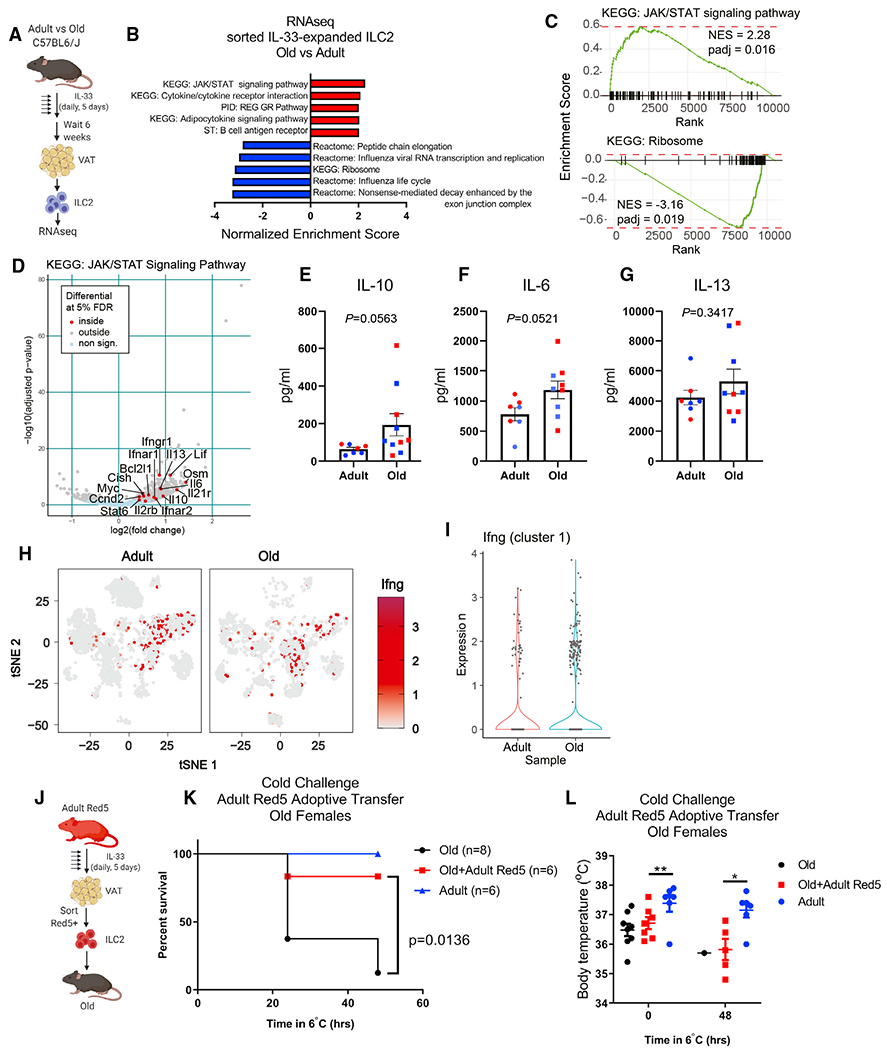
Aging impairs the integrated immunometabolic responses, which have evolved to maintain core body temperature in homeotherms to survive cold stress, infections, and dietary restriction. Adipose tissue inflammation regulates the thermogenic stress response, but how adipose tissue-resident cells instigate thermogenic failure in the aged are unknown. Here, we define alterations in the adipose-resident immune system and identify that type 2 innate lymphoid cells (ILC2s) are lost in aging. Restoration of ILC2 numbers in aged mice to levels seen in adults through IL-33 supplementation failed to rescue old mice from metabolic impairment and increased cold-induced lethality. Transcriptomic analyses revealed intrinsic defects in aged ILC2, and adoptive transfer of adult ILC2s are sufficient to protect old mice against cold. Thus, the functional defects in adipose ILC2s during aging drive thermogenic failure.
Keywords: IL-33; ILC2; adipose; aging; inflammation; metabolism; thermogenesis.
Copyright © 2021. Published by Elsevier Inc.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures

Comment in
-
Ageing immune cells reduce adipose tissue function.Nat Rev Endocrinol. 2021 Nov;17(11):638. doi: 10.1038/s41574-021-00574-2. Nat Rev Endocrinol. 2021. PMID: 34545230 No abstract available.
References
-
- Beretta E, Dhillon H, Kalra PS, and Kalra SP (2002). Central LIF gene therapy suppresses food intake, body weight, serum leptin and insulin for extended periods. Peptides 23, 975–984. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
