

Liolophura species discrimination with geographical distribution patterns and their divergence and expansion history on the northwestern Pacific coast
- PMID: 34475451
- PMCID: PMC8413323
- DOI: 10.1038/s41598-021-96823-5
Liolophura species discrimination with geographical distribution patterns and their divergence and expansion history on the northwestern Pacific coast
Abstract
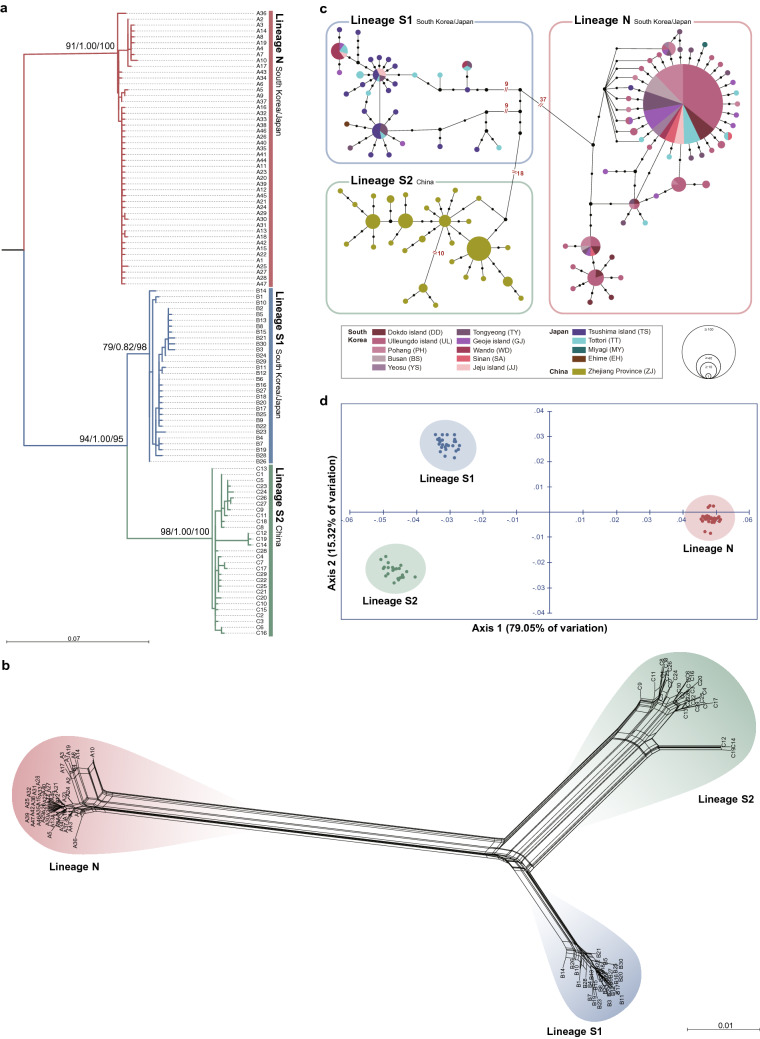
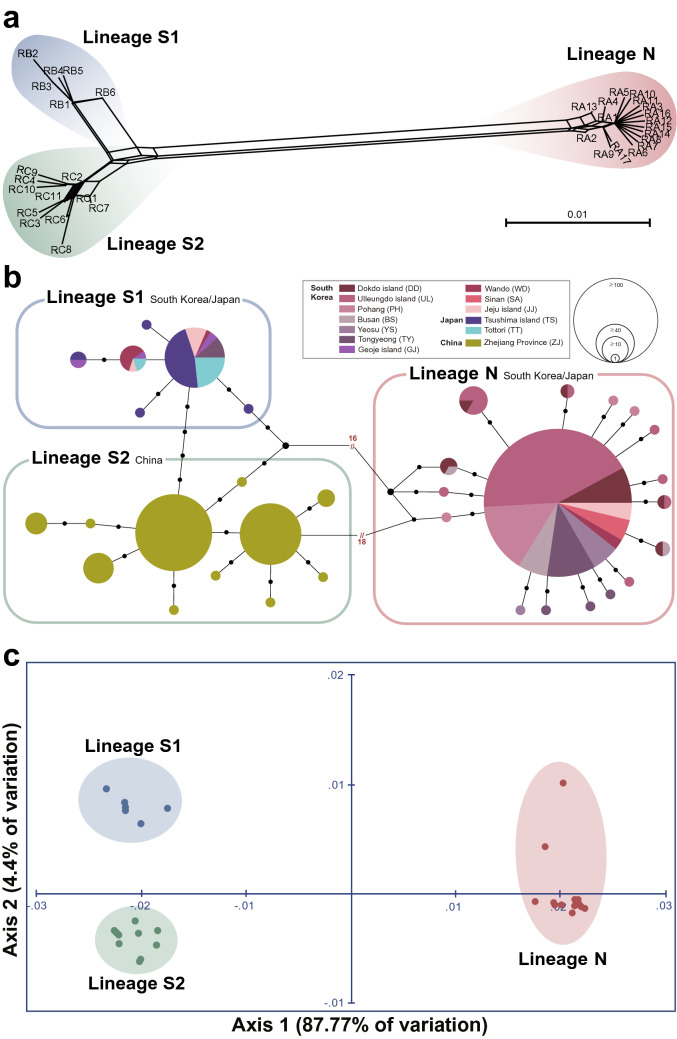
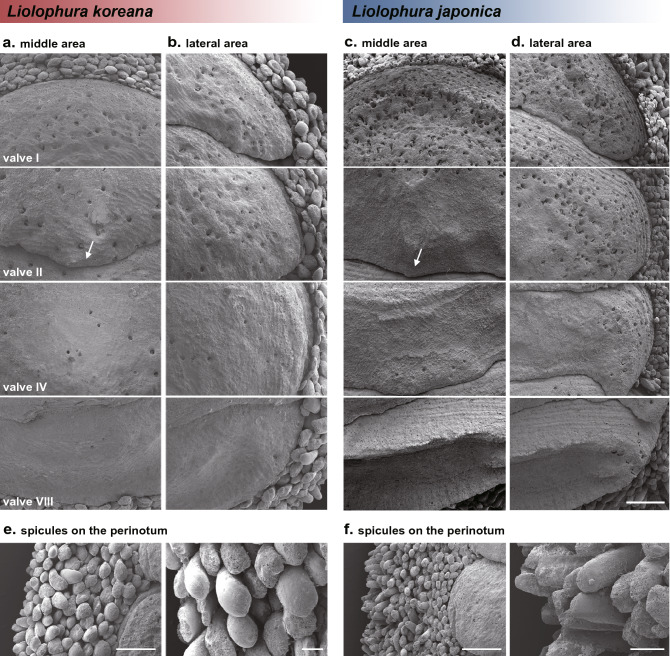
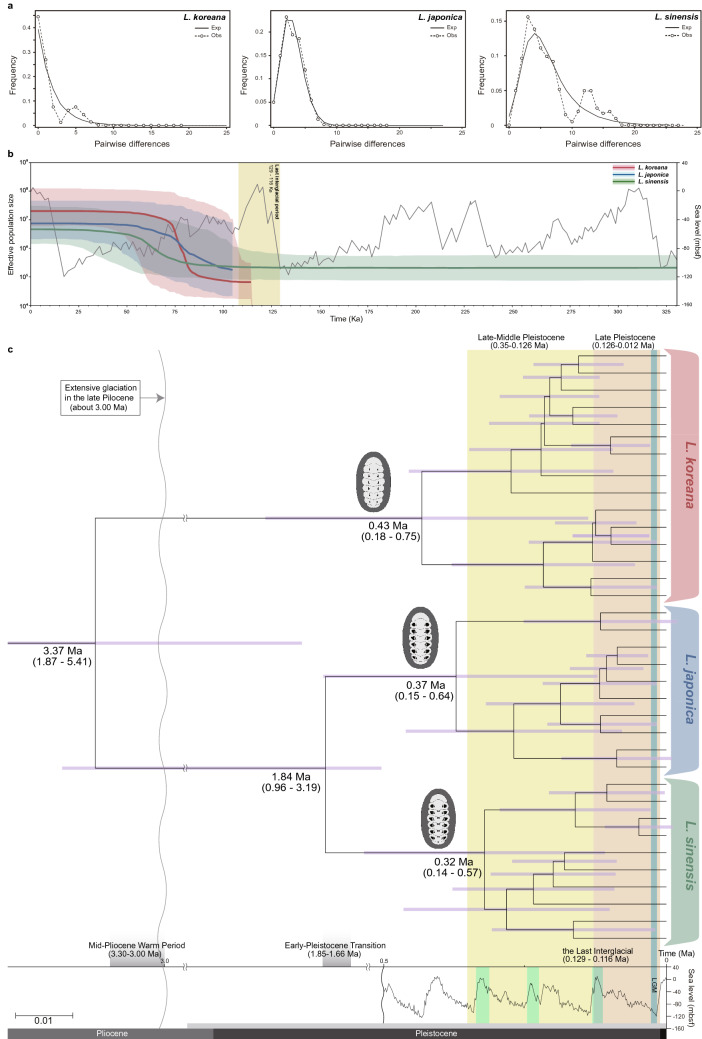
The chiton Liolophura japonica (Lischke 1873) is distributed in intertidal areas of the northwestern Pacific. Using COI and 16S rRNA, we found three genetic lineages, suggesting separation into three different species. Population genetic analyses, the two distinct COI barcoding gaps albeit one barcoding gap in the 16S rRNA, and phylogenetic relationships with a congeneric species supported this finding. We described L. koreana, sp. nov. over ca. 33°24' N (JJ), and L. sinensis, sp. nov. around ca. 27°02'-28°00' N (ZJ). We confirmed that these can be morphologically distinguished by lateral and dorsal black spots on the tegmentum and the shape of spicules on the perinotum. We also discuss species divergence during the Plio-Pleistocene, demographic expansions following the last interglacial age in the Pleistocene, and augmentation of COI haplotype diversity during the Pleistocene. Our study sheds light on the potential for COI in examining marine invertebrate species discrimination and distribution in the northwestern Pacific.
© 2021. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures




References
-
- Schwabe E. A catalogue of recent and fossil chitons (Mollusca: Polyplacophora) Addenda. Novapex. 2005;6:89–105.
-
- Sigwart JD. Morphological cladistic analysis as a model for character evaluation in primitive living chitons (Polyplacophora, Lepidopleurina) Am. Malacol. Bull. 2009;27:95–104. doi: 10.4003/006.027.0208. - DOI
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
