

Rewiring of human neurodevelopmental gene regulatory programs by human accelerated regions
- PMID: 34478631
- PMCID: PMC8542612
- DOI: 10.1016/j.neuron.2021.08.005
Rewiring of human neurodevelopmental gene regulatory programs by human accelerated regions
Abstract
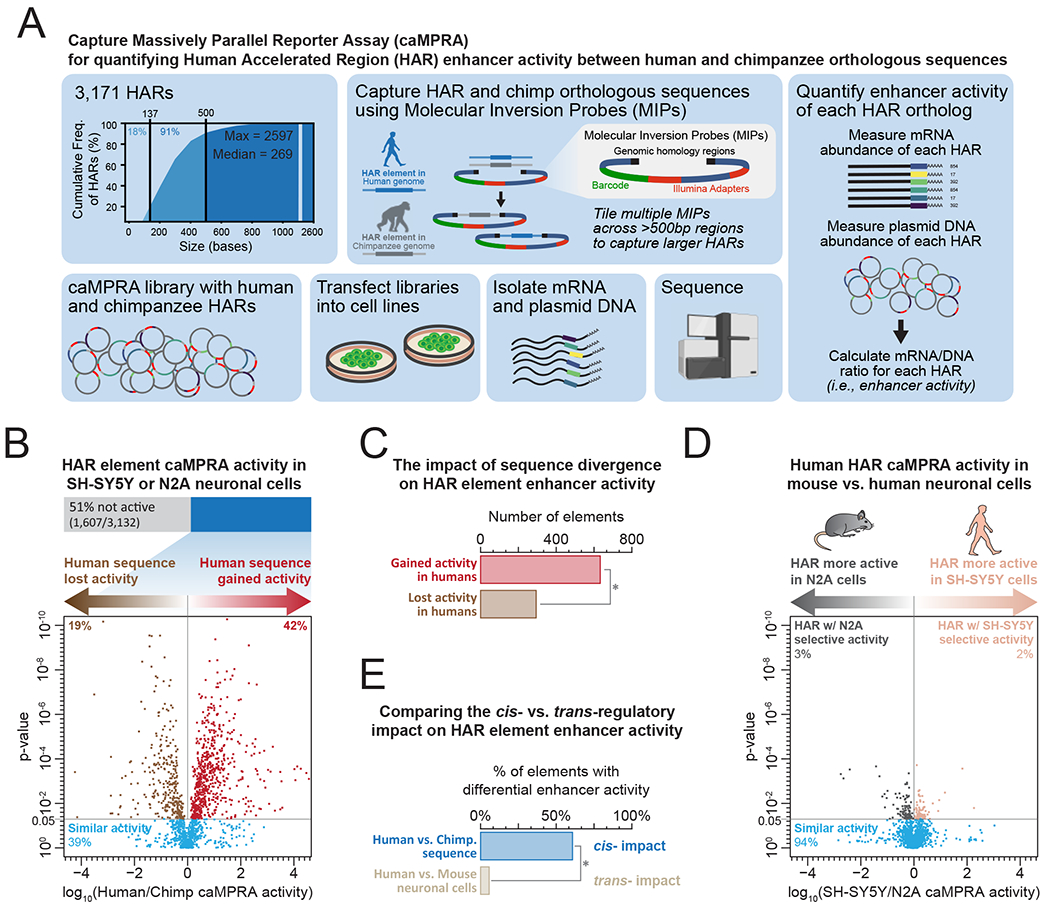
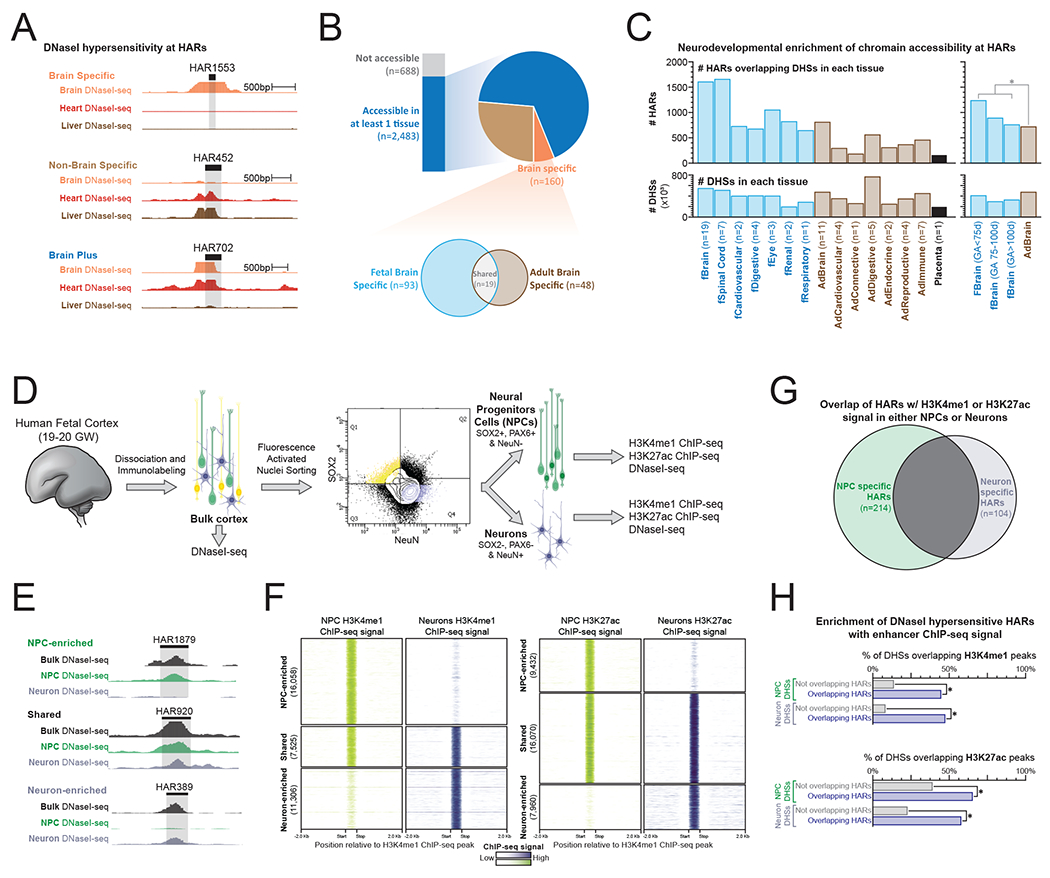
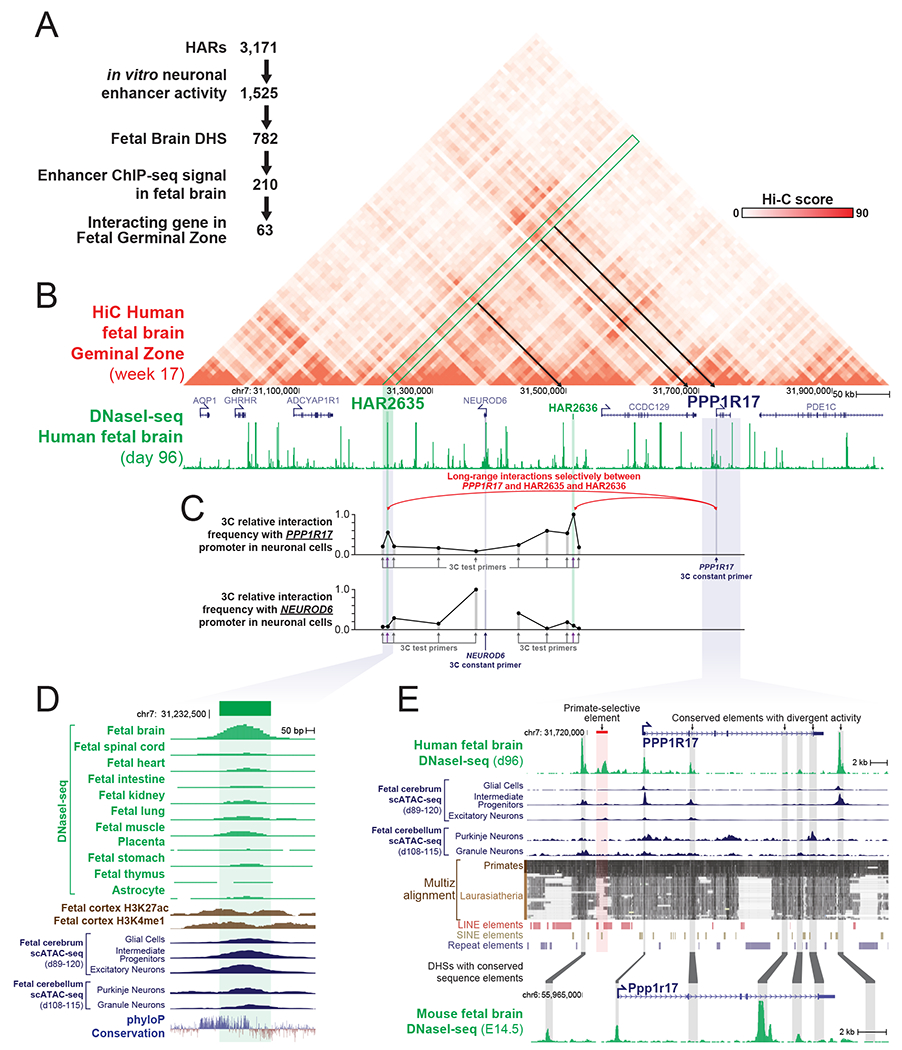
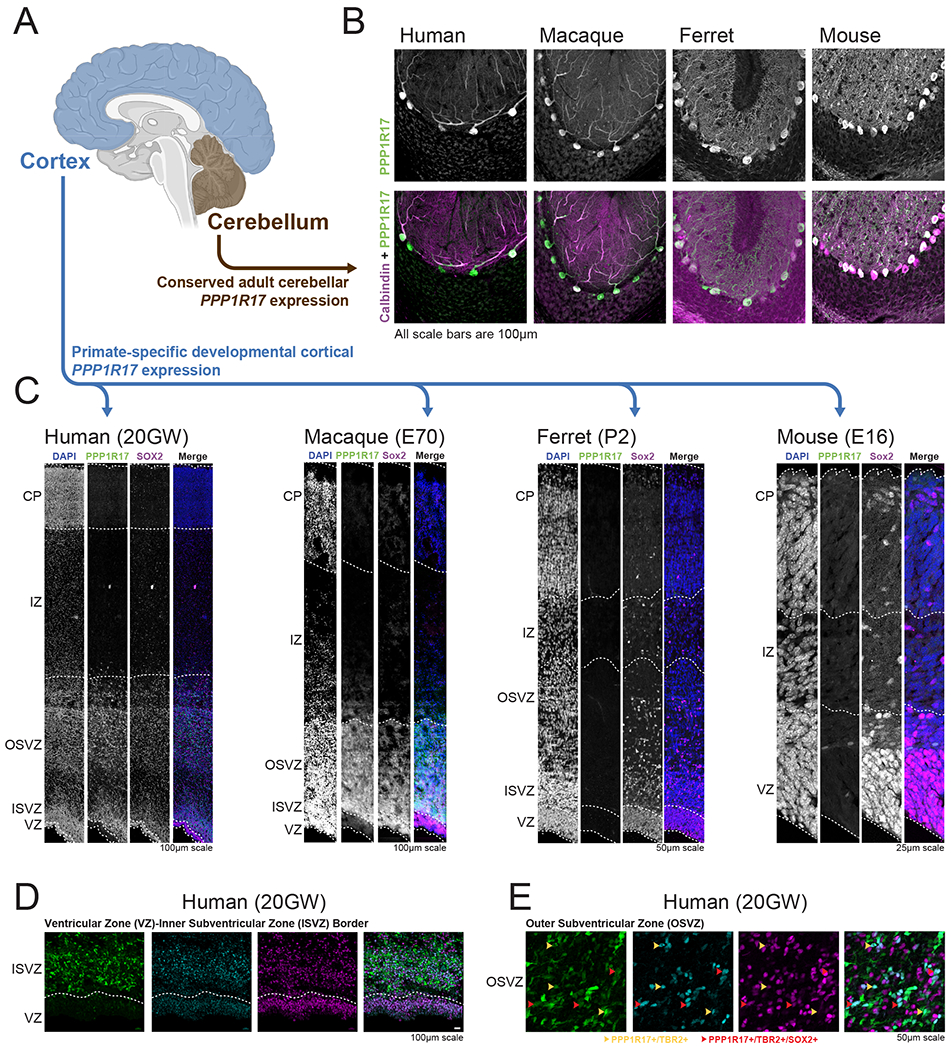
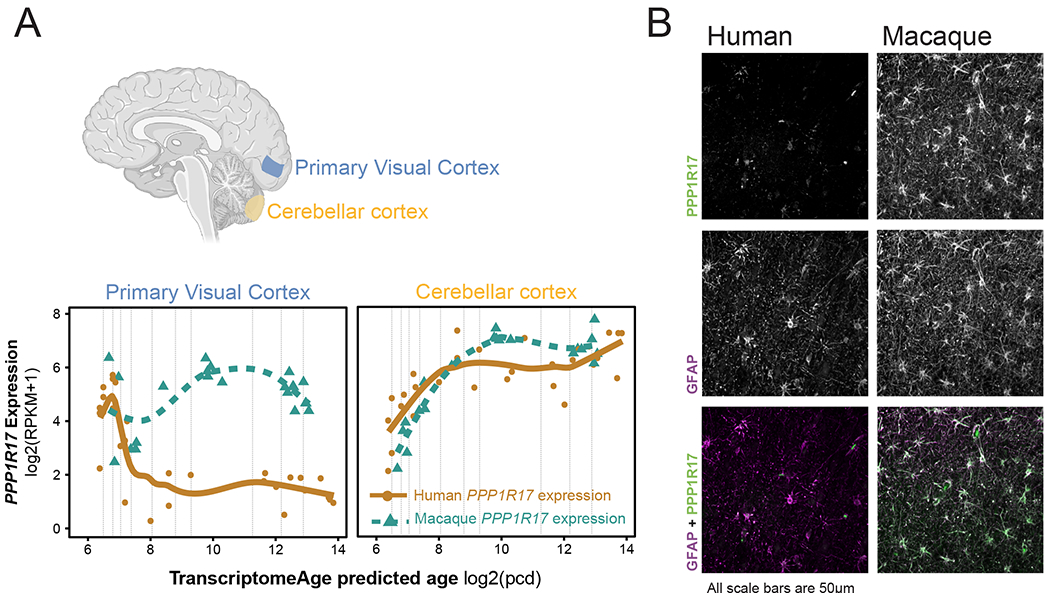
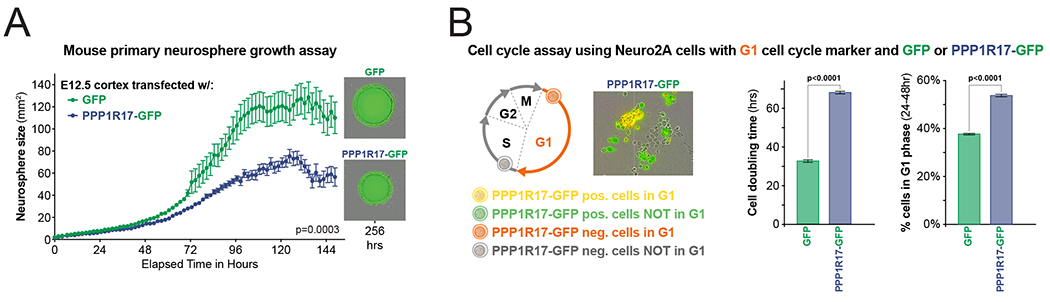
Human accelerated regions (HARs) are the fastest-evolving regions of the human genome, and many are hypothesized to function as regulatory elements that drive human-specific gene regulatory programs. We interrogate the in vitro enhancer activity and in vivo epigenetic landscape of more than 3,100 HARs during human neurodevelopment, demonstrating that many HARs appear to act as neurodevelopmental enhancers and that sequence divergence at HARs has largely augmented their neuronal enhancer activity. Furthermore, we demonstrate PPP1R17 to be a putative HAR-regulated gene that has undergone remarkable rewiring of its cell type and developmental expression patterns between non-primates and primates and between non-human primates and humans. Finally, we show that PPP1R17 slows neural progenitor cell cycle progression, paralleling the cell cycle length increase seen predominantly in primate and especially human neurodevelopment. Our findings establish HARs as key components in rewiring human-specific neurodevelopmental gene regulatory programs and provide an integrated resource to study enhancer activity of specific HARs.
Keywords: HARs; cerebral cortex; enhancers; evolution; human accelerated regions; neurodevelopment.
Copyright © 2021 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
Comment in
-
Uncovering the HARbingers of human brain evolution.Neuron. 2021 Oct 20;109(20):3231-3233. doi: 10.1016/j.neuron.2021.09.022. Neuron. 2021. PMID: 34672980
References
-
- Betizeau M, Cortay V, Patti D, Pfister S, Gautier E, Bellemin-Ménard A, Afanassieff M, Huissoud C, Douglas RJ, Kennedy H, et al. (2013). Precursor Diversity and Complexity of Lineage Relationships in the Outer Subventricular Zone of the Primate. Neuron 80, 442–457. - PubMed
Publication types
MeSH terms
Grants and funding
- P50 HD105351/HD/NICHD NIH HHS/United States
- T32 GM007753/GM/NIGMS NIH HHS/United States
- R01 NS095654/NS/NINDS NIH HHS/United States
- DP5 OD029630/OD/NIH HHS/United States
- HHMI/Howard Hughes Medical Institute/United States
- R01 NS115965/NS/NINDS NIH HHS/United States
- U54 HD090255/HD/NICHD NIH HHS/United States
- T32 GM007748/GM/NIGMS NIH HHS/United States
- U01 MH116488/MH/NIMH NIH HHS/United States
- P50 MH106934/MH/NIMH NIH HHS/United States
- S10 RR028832/RR/NCRR NIH HHS/United States
- R01 NS032457/NS/NINDS NIH HHS/United States
- K00 CA222750/CA/NCI NIH HHS/United States
- T32 MH020017/MH/NIMH NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
