

Mechanosensory input during circuit formation shapes Drosophila motor behavior through patterned spontaneous network activity
- PMID: 34478644
- PMCID: PMC8665011
- DOI: 10.1016/j.cub.2021.08.022
Mechanosensory input during circuit formation shapes Drosophila motor behavior through patterned spontaneous network activity
Abstract
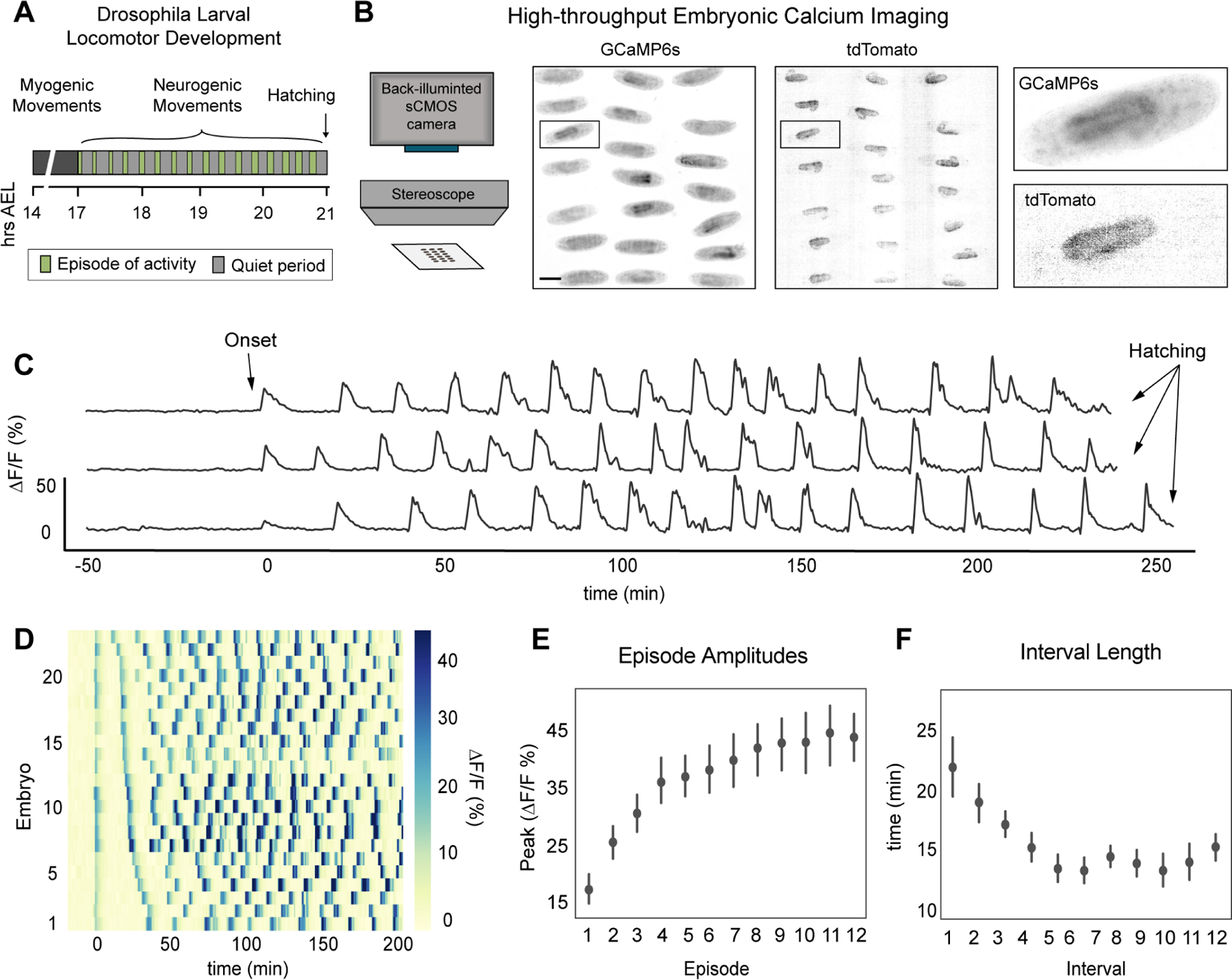
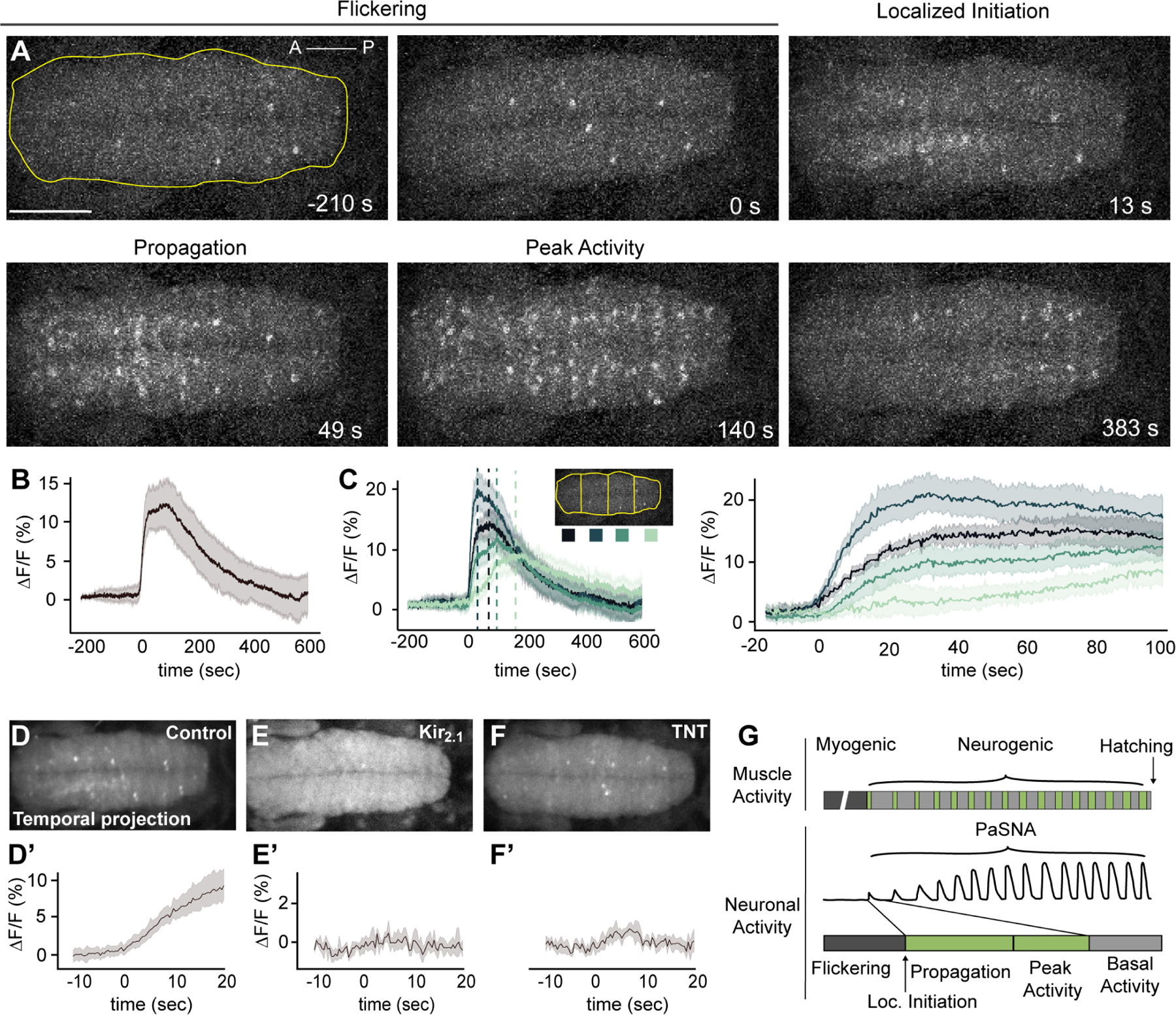
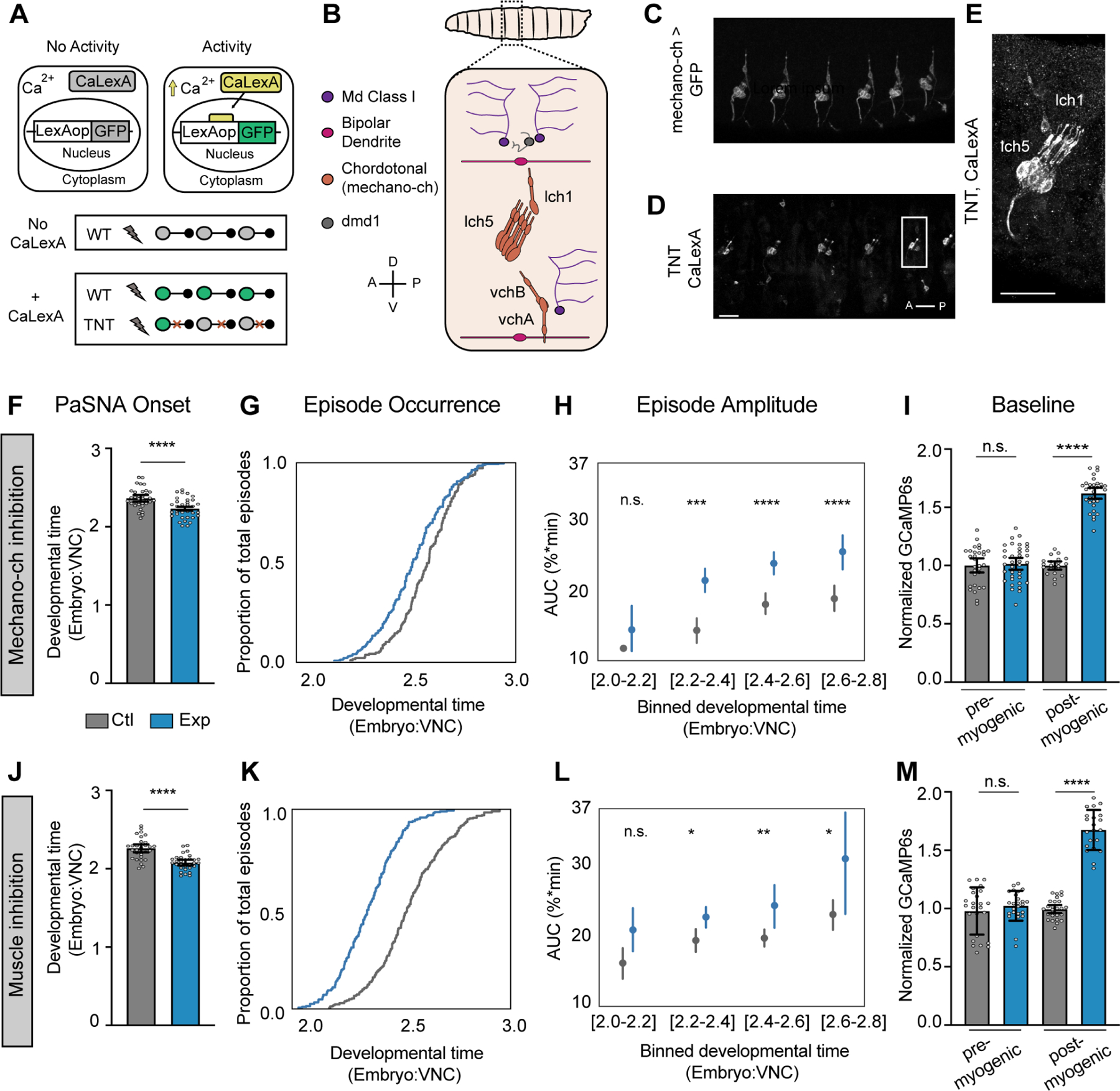
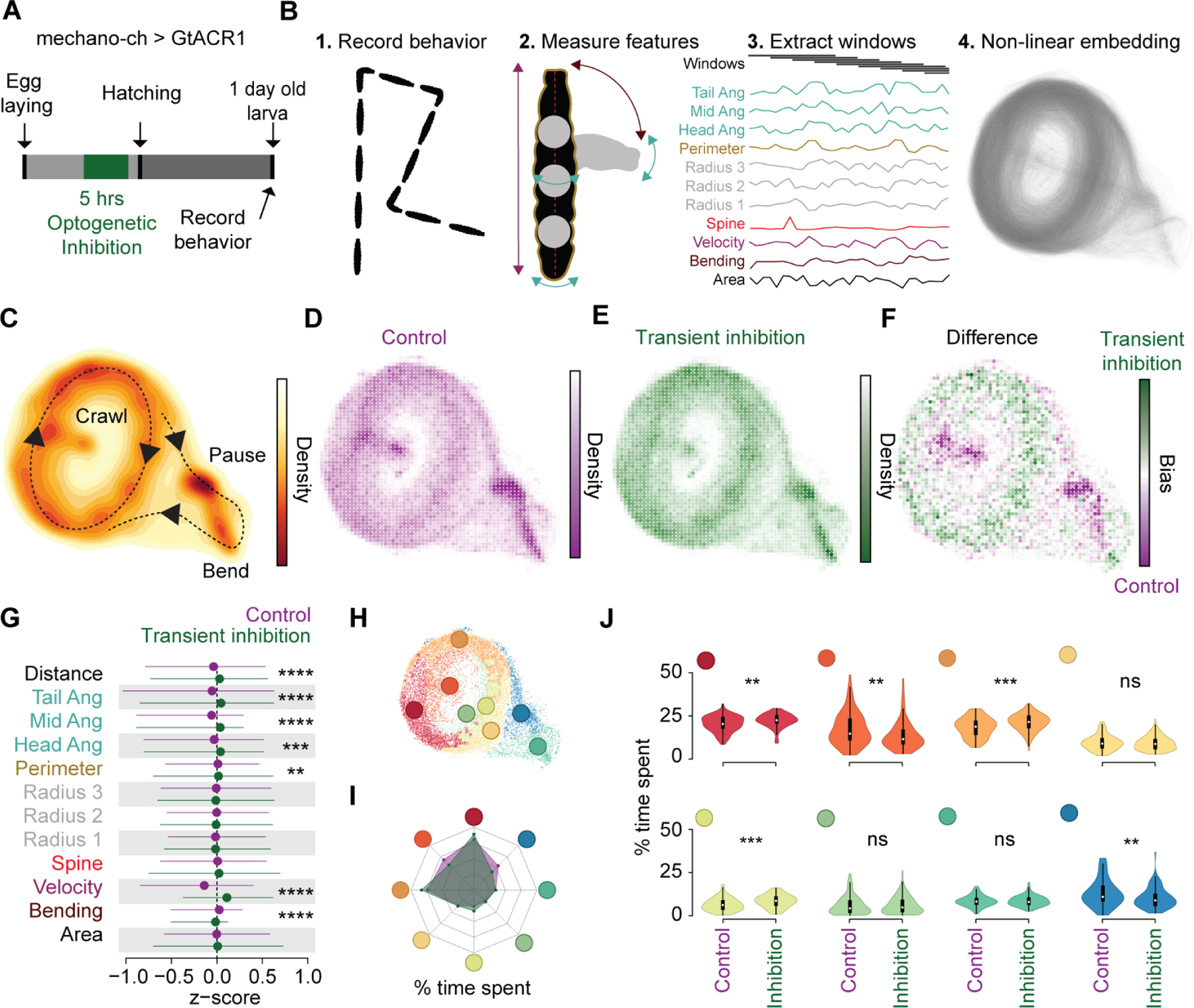
Neural activity sculpts circuit wiring in many animals. In vertebrates, patterned spontaneous network activity (PaSNA) generates sensory maps and establishes local circuits.1-3 However, it remains unclear how PaSNA might shape neuronal circuits and behavior in invertebrates. Previous work in the developing Drosophila embryo discovered intrinsic muscle activity that did not require synaptic transmission, and hence was myogenic, preceding PaSNA.4-6 These studies, however, monitored muscle movement, not neural activity, and were therefore unable to observe how myogenic activity might relate to subsequent neural network engagement. Here we use calcium imaging to directly record neural activity and characterize the emergence of PaSNA. We demonstrate that the spatiotemporal properties of PaSNA are highly stereotyped across embryos, arguing for genetic programming. Neural activity begins well before it becomes patterned, emerging during the myogenic stage. Remarkably, inhibition of mechanosensory input, as well as inhibition of muscle contractions, results in premature and excessive PaSNA, demonstrating that muscle movement serves as a brake on this process. Finally, transient mechanosensory inhibition during PaSNA, followed by quantitative modeling of larval behavior, shows that mechanosensory modulation during development is required for proper larval foraging. This work provides a foundation for using the Drosophila embryo to study the role of PaSNA in circuit formation, provides mechanistic insight into how PaSNA is entrained by motor activity, and demonstrates that spontaneous network activity is essential for locomotor behavior. These studies argue that sensory feedback during the earliest stages of circuit formation can sculpt locomotor behaviors through innate motor learning.
Keywords: Drosophila embryo; behavioral development; development of locomotor behavior; nervous system development; neural circuit wiring; spontaneous network activity.
Copyright © 2021 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
Comment in
-
Neural development: The role of spontaneous activity.Curr Biol. 2021 Dec 6;31(23):R1513-R1515. doi: 10.1016/j.cub.2021.10.026. Curr Biol. 2021. PMID: 34875237
References
-
- Goodman CS, and Shatz CJ (1993). Developmental mechanisms that generate precise patterns of neuronal connectivity. Cell 72, 77–98. - PubMed
-
- Pereanu W, Spindler S, Im E, Buu N, and Hartenstein V. (2007). The Emergence of Patterned Movement During Late Embryogenesis of Drosophila. Dev Neurobiology, 1669–1685. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
