

An Integrated Transcriptomics and Proteomics Analysis Implicates lncRNA MALAT1 in the Regulation of Lipid Metabolism
- PMID: 34478876
- PMCID: PMC8479834
- DOI: 10.1016/j.mcpro.2021.100141
An Integrated Transcriptomics and Proteomics Analysis Implicates lncRNA MALAT1 in the Regulation of Lipid Metabolism
Abstract
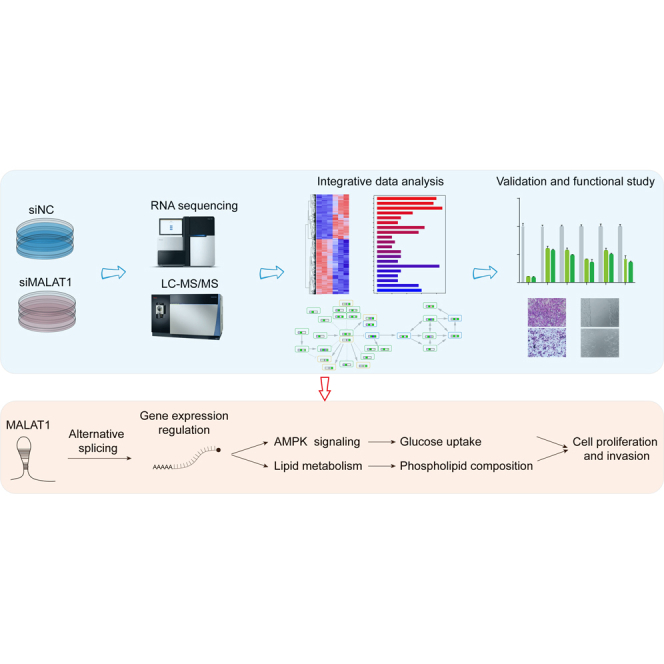
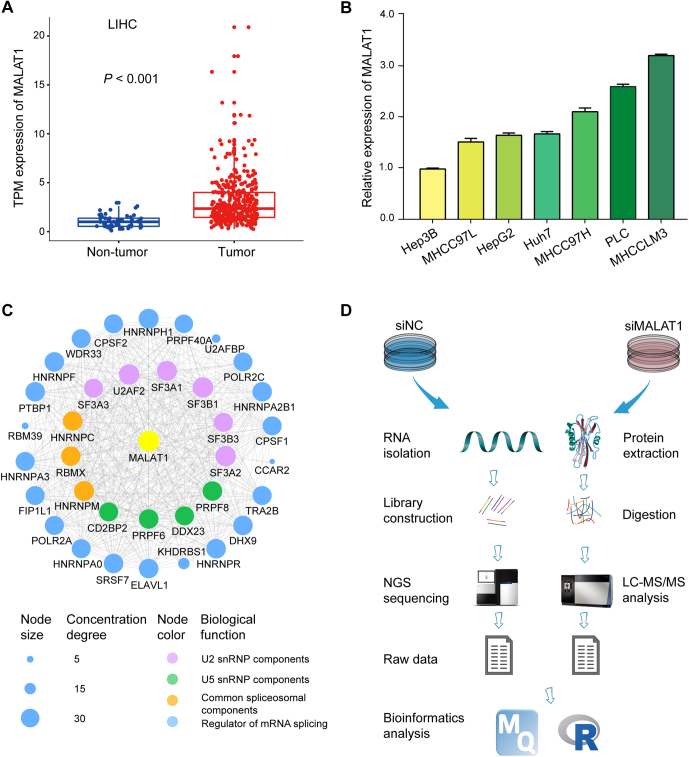
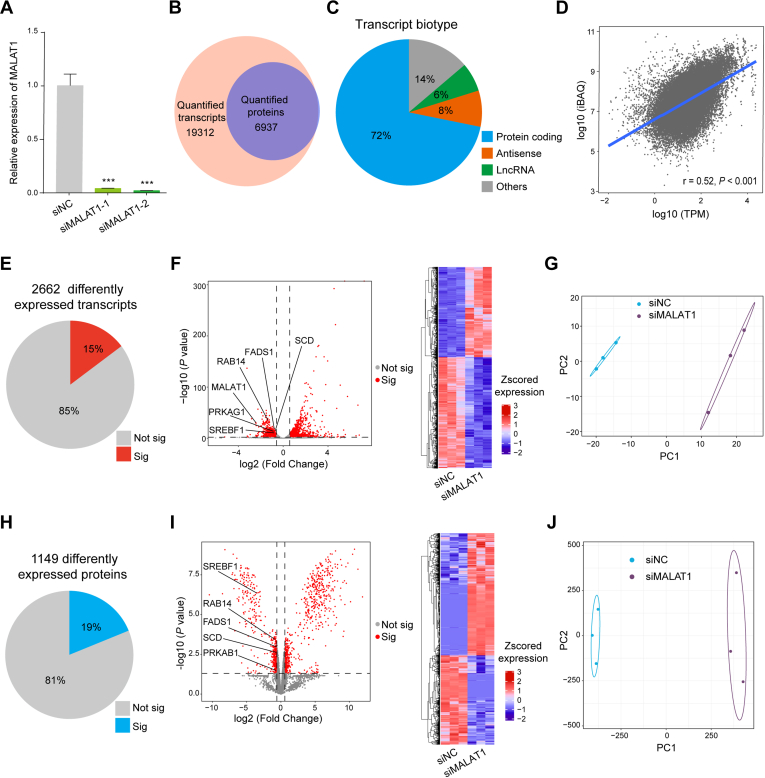
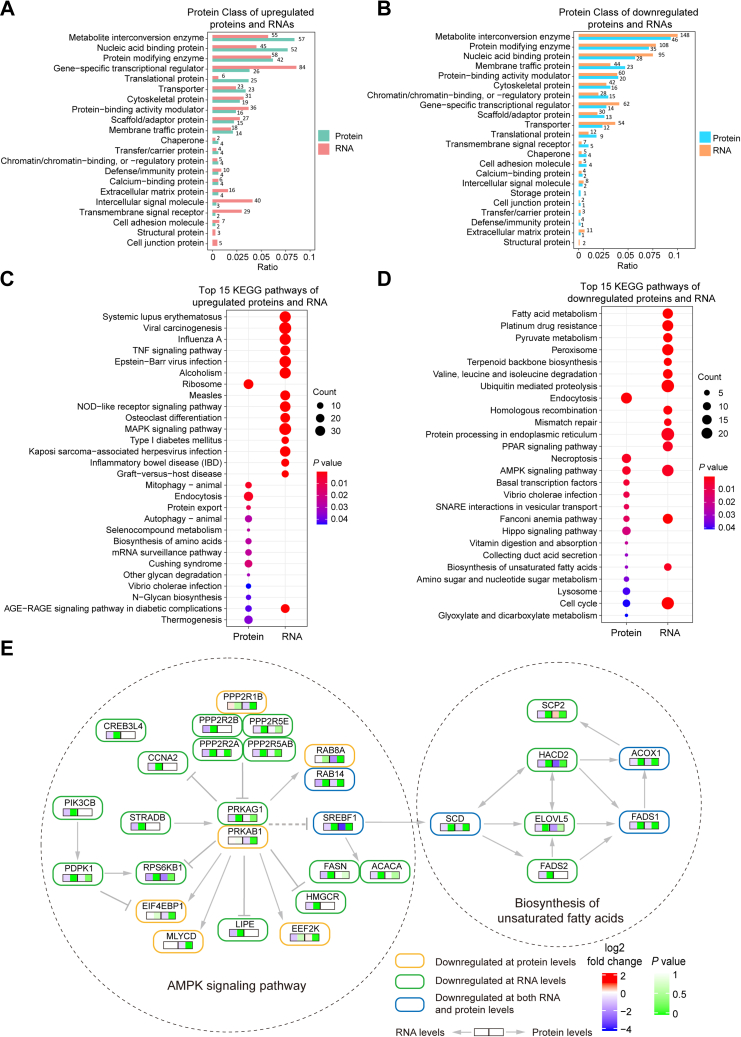
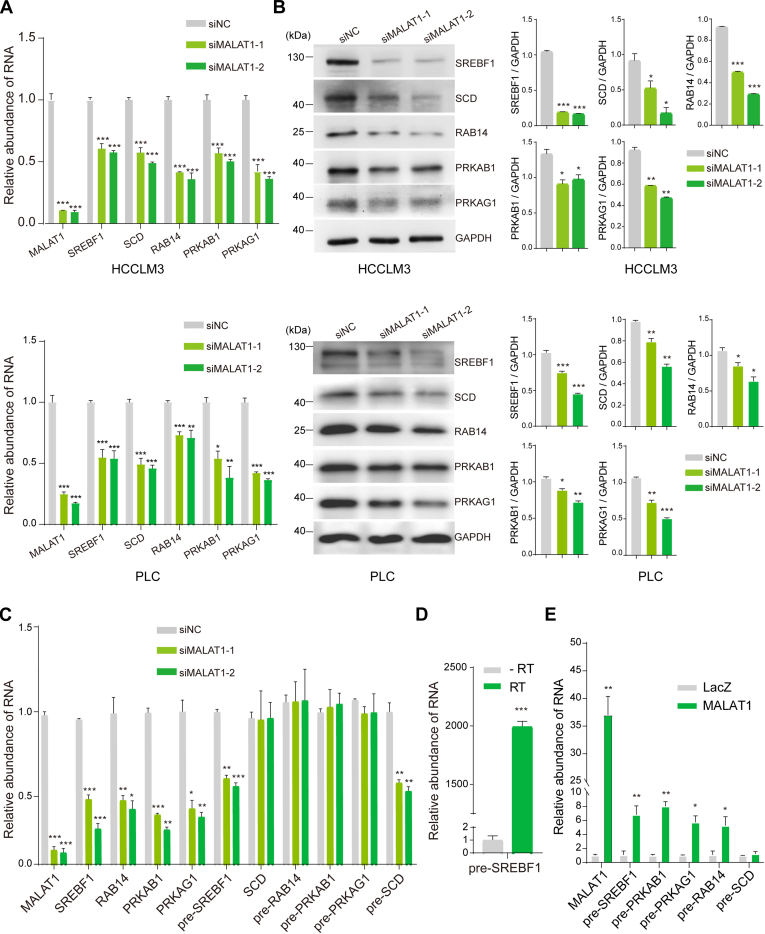
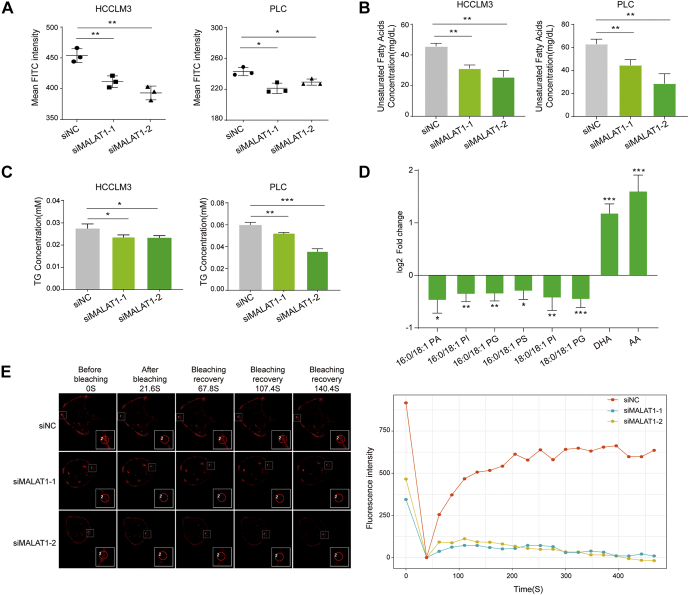
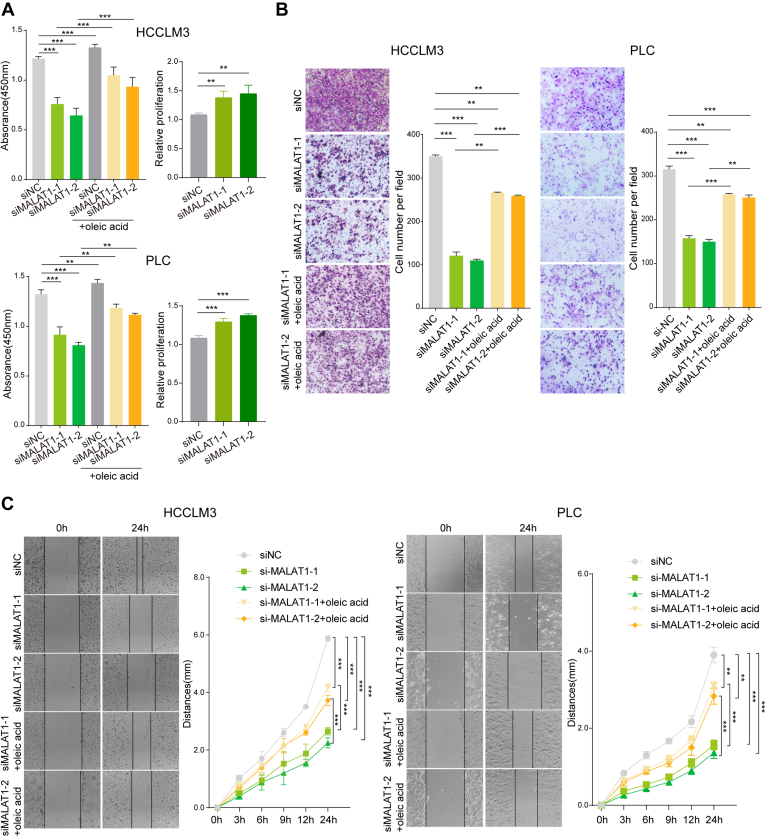
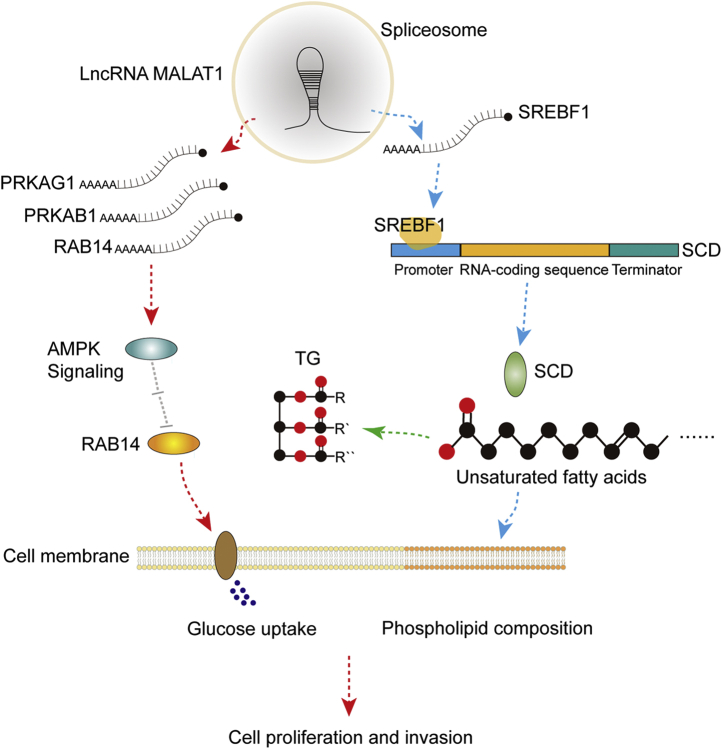
Long noncoding RNA metastasis-associated lung adenocarcinoma transcript 1 (MALAT1) is upregulated in various cancers, and its overexpression is associated with tumor growth and metastasis. MALAT1 has been recognized as a key player in the regulation of RNA splicing and transcription; however, the landscape of gene expression regulated by MALAT1 remains unclear. In this study, we employed an integrated transcriptomics and proteomics strategy to characterize the alterations in gene expression induced by MALAT1 knockdown in hepatocellular carcinoma (HCC) cells and identified 2662 differentially expressed transcripts and 1149 differentially expressed proteins. Interestingly, downregulation of MALAT1 reduced the abundances of multiple genes in the AMP-activated protein kinase (AMPK) signaling and biosynthesis of unsaturated fatty acids pathways. Further investigation showed that MALAT1 knockdown inhibited glucose uptake and lipogenesis by reducing the expression levels of these lipid metabolism related genes, which contributes to the oncogenic role of MALAT1 in tumor cell proliferation and invasion. This study uncovers the function of MALAT1 in the modulation of cancer lipid metabolism, reveals the underlying molecular mechanism, and further supports the potential therapeutic opportunities for targeting MALAT1 in HCC treatment.
Keywords: hepatocellular carcinoma; lipid metabolism; long noncoding RNA; proteomics; transcriptomics.
Copyright © 2021 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Conflict of interest The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Figures
References
-
- Robey R.B., Weisz J., Kuemmerle N.B., Salzberg A.C., Berg A., Brown D.G., Kubik L., Palorini R., Al-Mulla F., Al-Temaimi R., Colacci A., Mondello C., Raju J., Woodrick J., Scovassi A.I. Metabolic reprogramming and dysregulated metabolism: Cause, consequence and/or enabler of environmental carcinogenesis? Carcinogenesis. 2015;36:S203–S231. - PMC - PubMed
-
- Budhu A., Roessler S., Zhao X., Yu Z., Forgues M., Ji J., Karoly E., Qin L.X., Ye Q.H., Jia H.L., Fan J., Sun H.C., Tang Z.Y., Wang X.W. Integrated metabolite and gene expression profiles identify lipid biomarkers associated with progression of hepatocellular carcinoma and patient outcomes. Gastroenterology. 2013;144:1066–1075.e1. - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
