Genetic Mechanism for the Cyclostome Cerebellar Neurons Reveals Early Evolution of the Vertebrate Cerebellum
- PMID: 34485287
- PMCID: PMC8416312
- DOI: 10.3389/fcell.2021.700860
Genetic Mechanism for the Cyclostome Cerebellar Neurons Reveals Early Evolution of the Vertebrate Cerebellum
Abstract
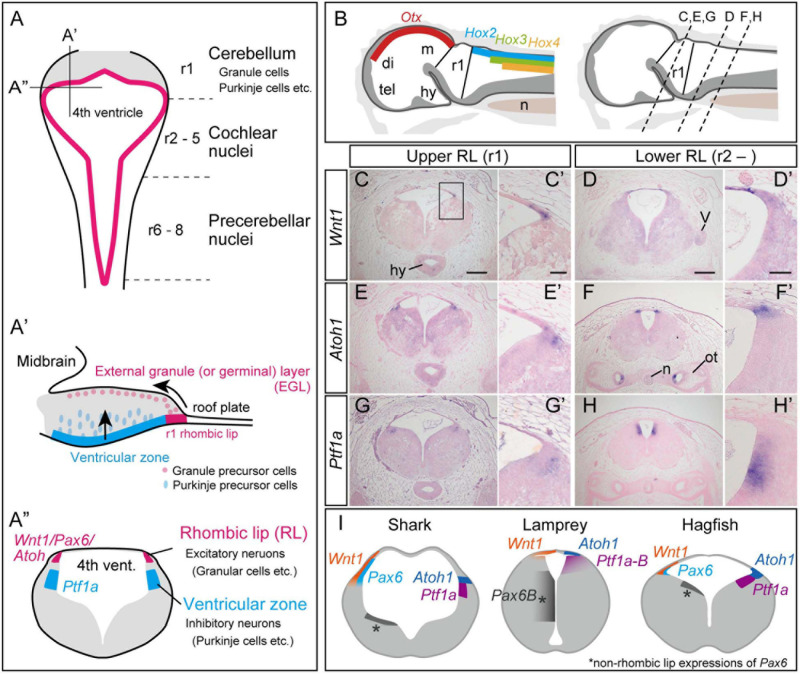
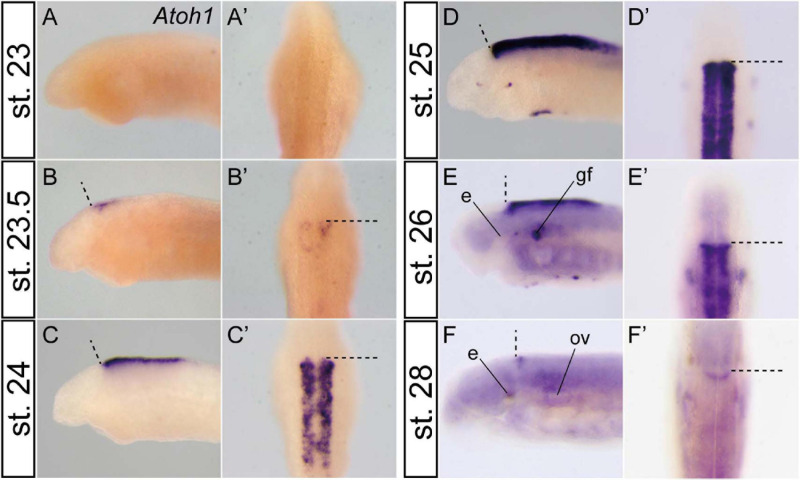
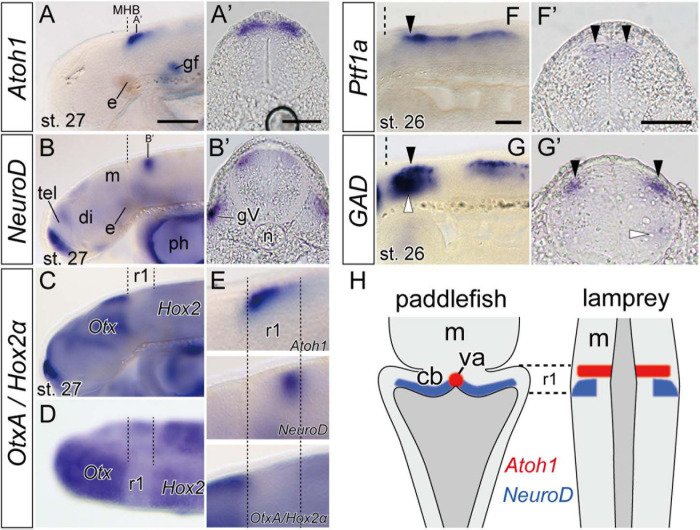
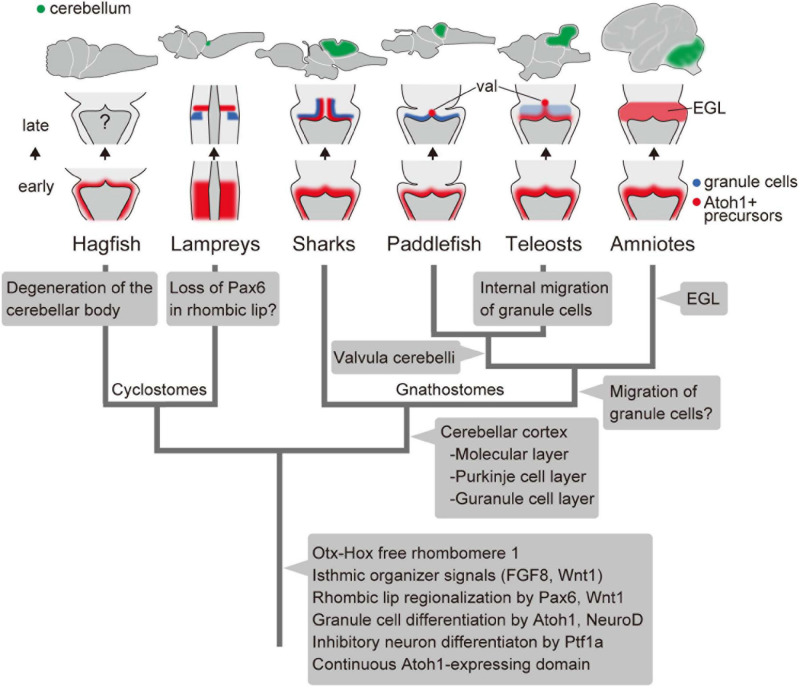
The vertebrate cerebellum arises at the dorsal part of rhombomere 1, induced by signals from the isthmic organizer. Two major cerebellar neuronal subtypes, granule cells (excitatory) and Purkinje cells (inhibitory), are generated from the anterior rhombic lip and the ventricular zone, respectively. This regionalization and the way it develops are shared in all extant jawed vertebrates (gnathostomes). However, very little is known about early evolution of the cerebellum. The lamprey, an extant jawless vertebrate lineage or cyclostome, possesses an undifferentiated, plate-like cerebellum, whereas the hagfish, another cyclostome lineage, is thought to lack a cerebellum proper. In this study, we found that hagfish Atoh1 and Wnt1 genes are co-expressed in the rhombic lip, and Ptf1a is expressed ventrally to them, confirming the existence of r1's rhombic lip and the ventricular zone in cyclostomes. In later stages, lamprey Atoh1 is downregulated in the posterior r1, in which the NeuroD increases, similar to the differentiation process of cerebellar granule cells in gnathostomes. Also, a continuous Atoh1-positive domain in the rostral r1 is reminiscent of the primordium of valvula cerebelli of ray-finned fishes. Lastly, we detected a GAD-positive domain adjacent to the Ptf1a-positive ventricular zone in lampreys, suggesting that the Ptf1a-positive cells differentiate into some GABAergic inhibitory neurons such as Purkinje and other inhibitory neurons like in gnathostomes. Altogether, we conclude that the ancestral genetic programs for the formation of a distinct cerebellum were established in the last common ancestor of vertebrates.
Keywords: Purkinje cells; cerebellum evolution; cyclostome; evolutionary developmental biology (EvoDevo); granule cells of cerebellum; hagfish; lamprey; rhombic lip.
Copyright © 2021 Sugahara, Pascual-Anaya, Kuraku, Kuratani and Murakami.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures