

In vivo deep network tracing reveals phosphofructokinase-mediated coordination of biosynthetic pathway activity in the myocardium
- PMID: 34487754
- PMCID: PMC8766935
- DOI: 10.1016/j.yjmcc.2021.08.013
In vivo deep network tracing reveals phosphofructokinase-mediated coordination of biosynthetic pathway activity in the myocardium
Abstract
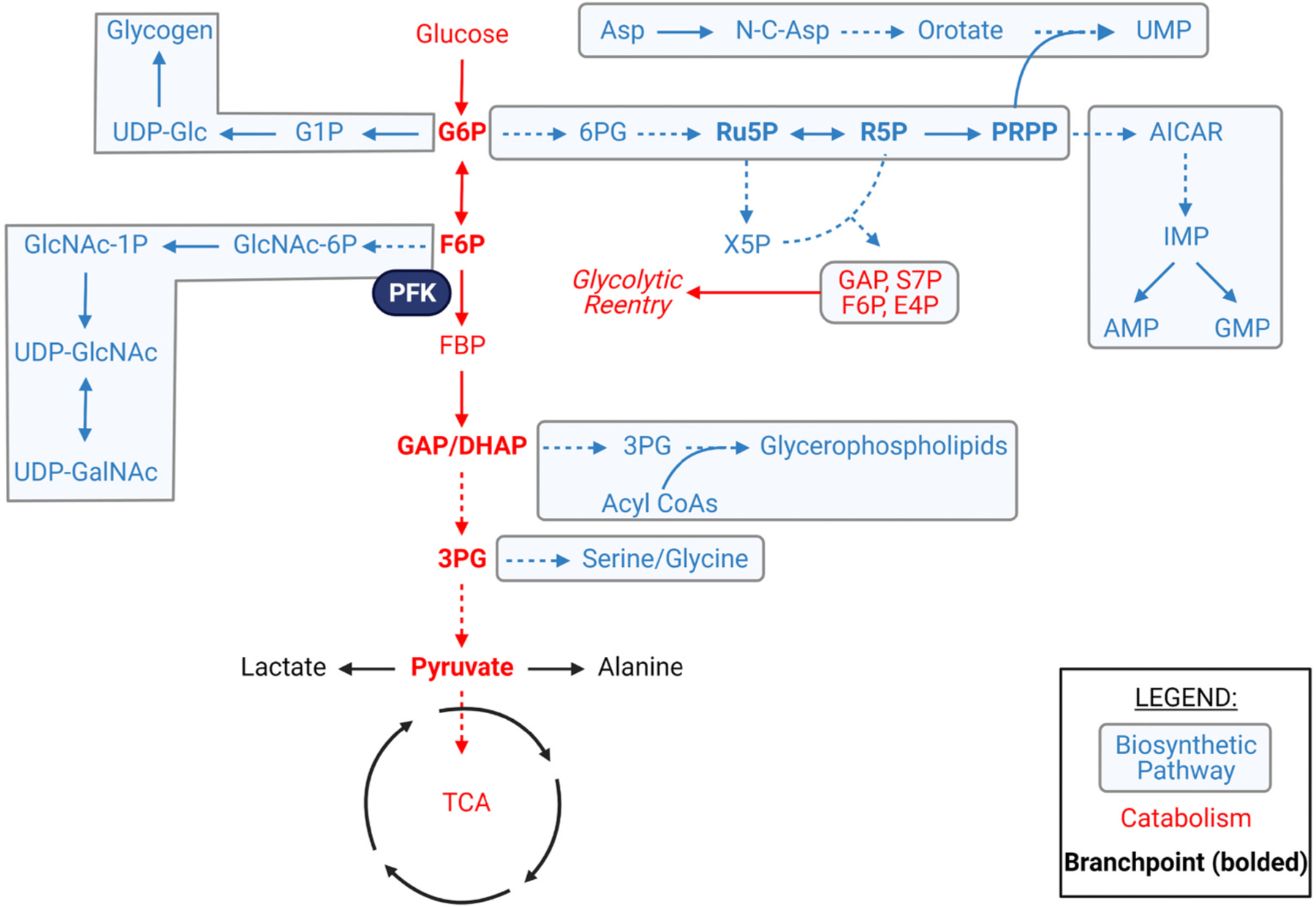
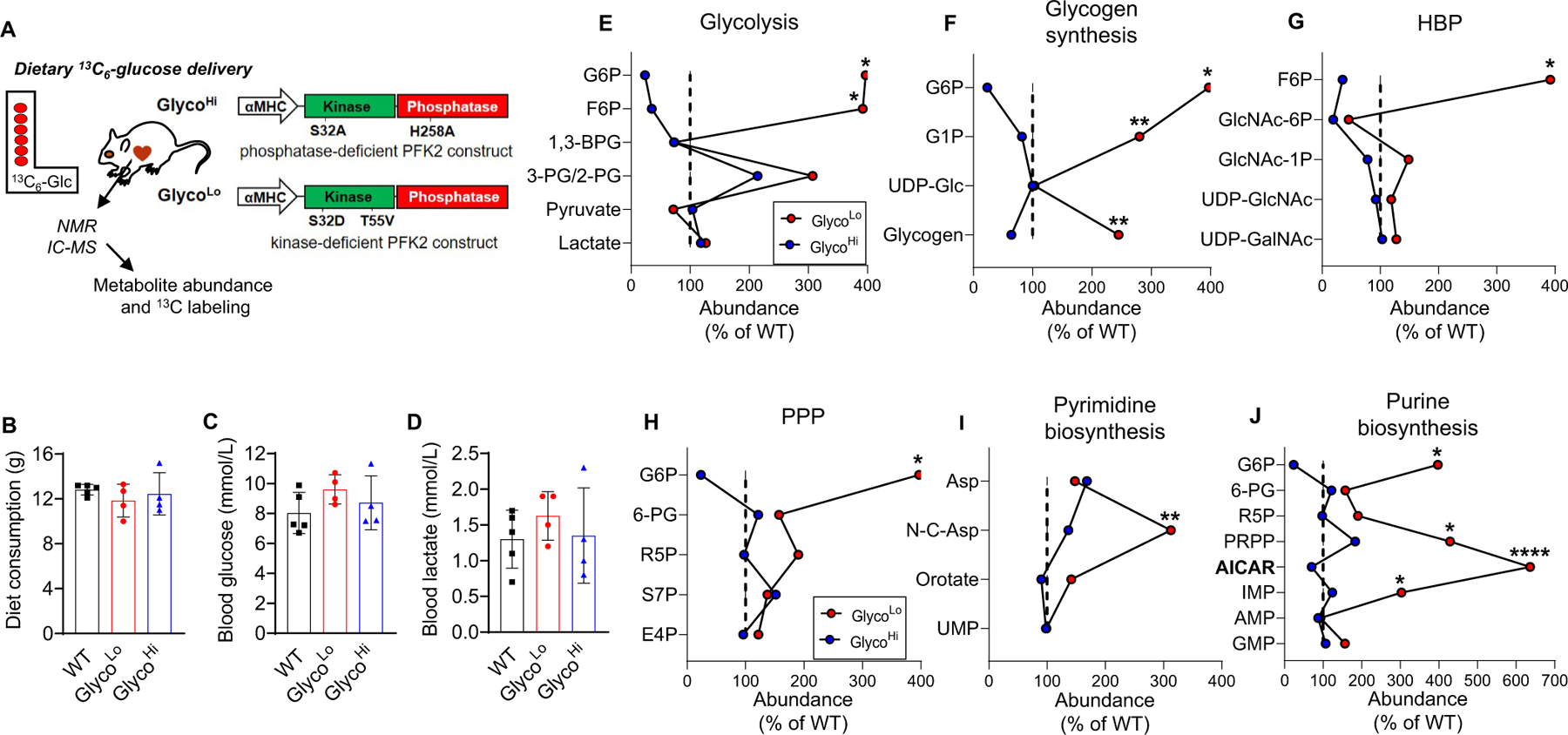
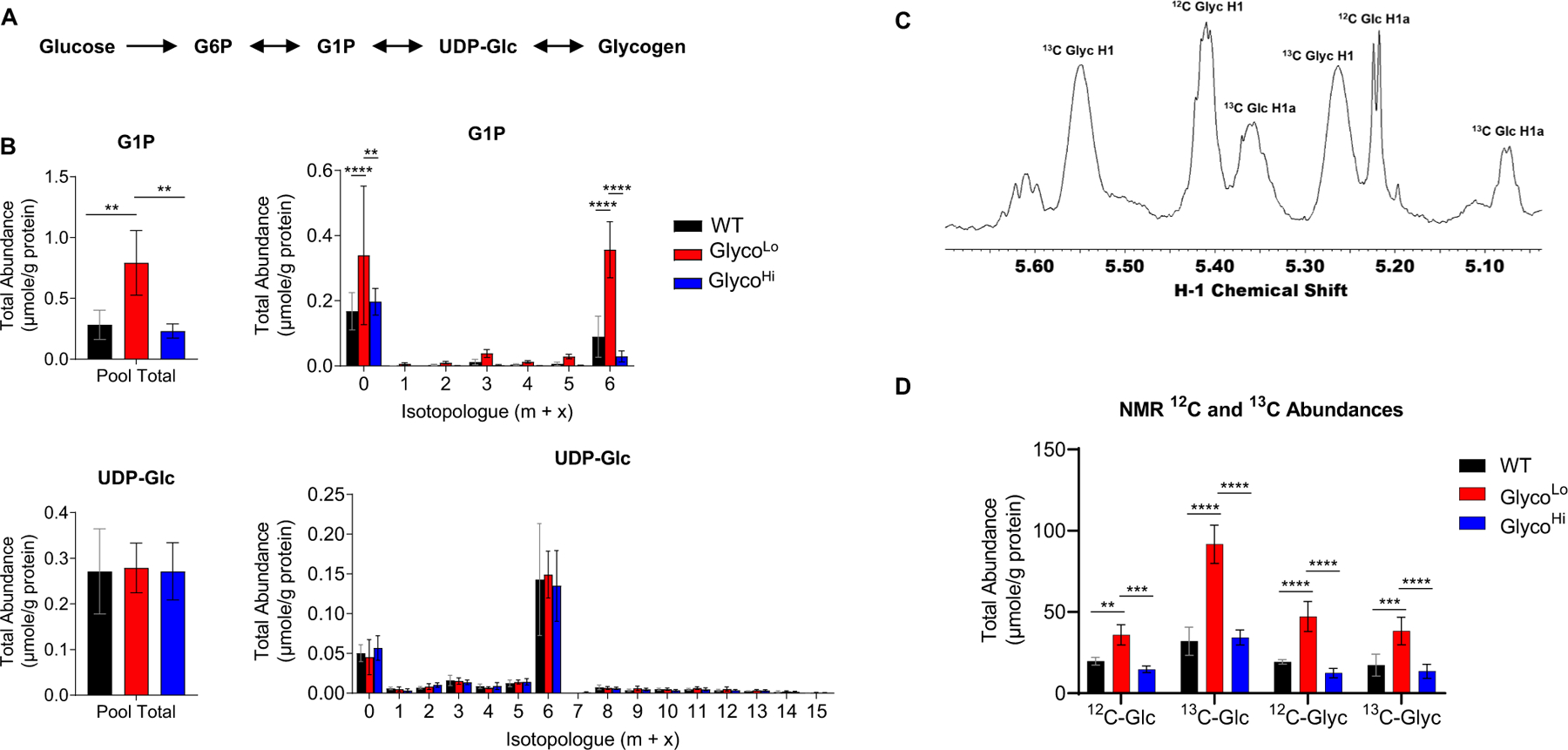
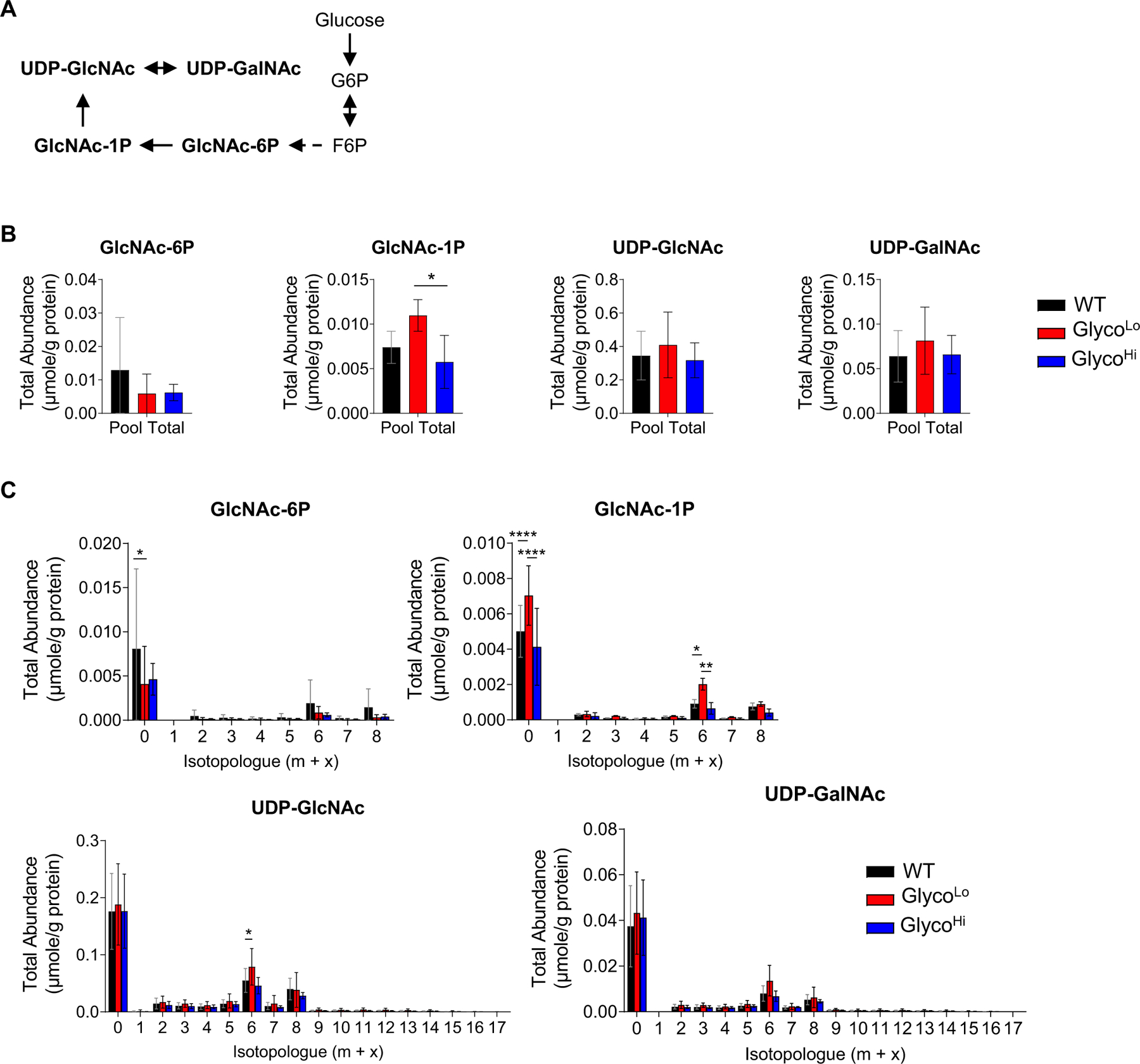
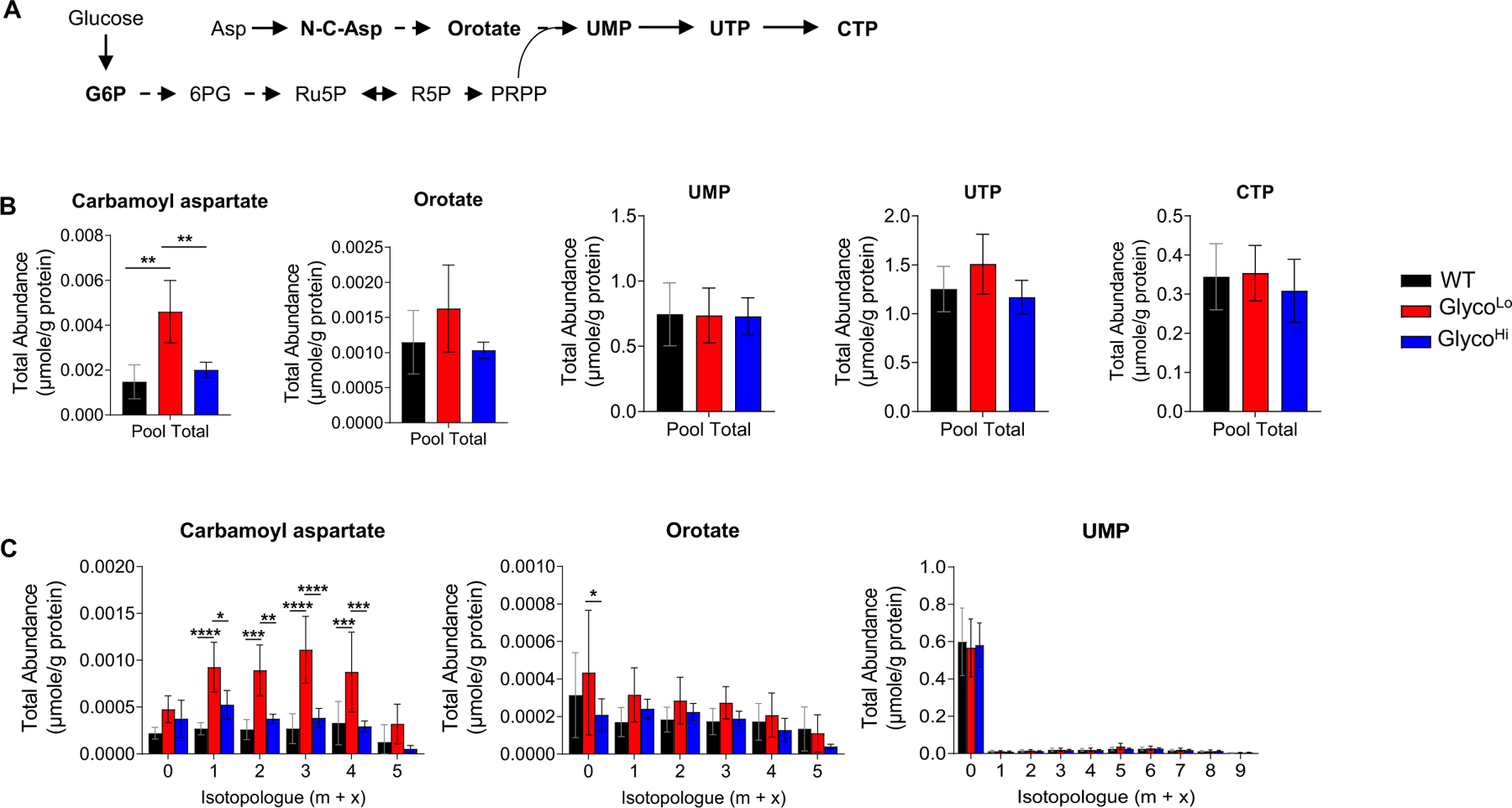
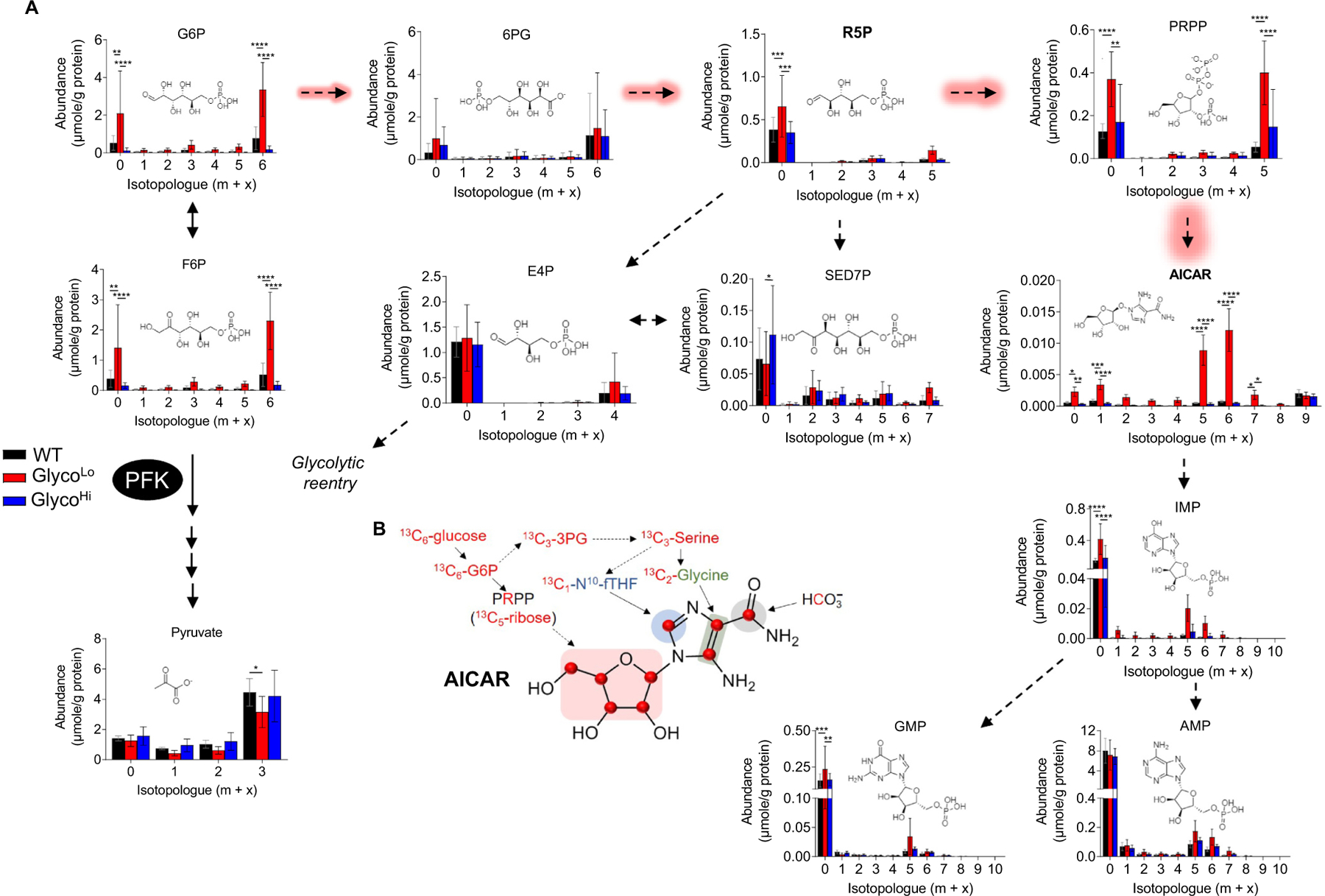
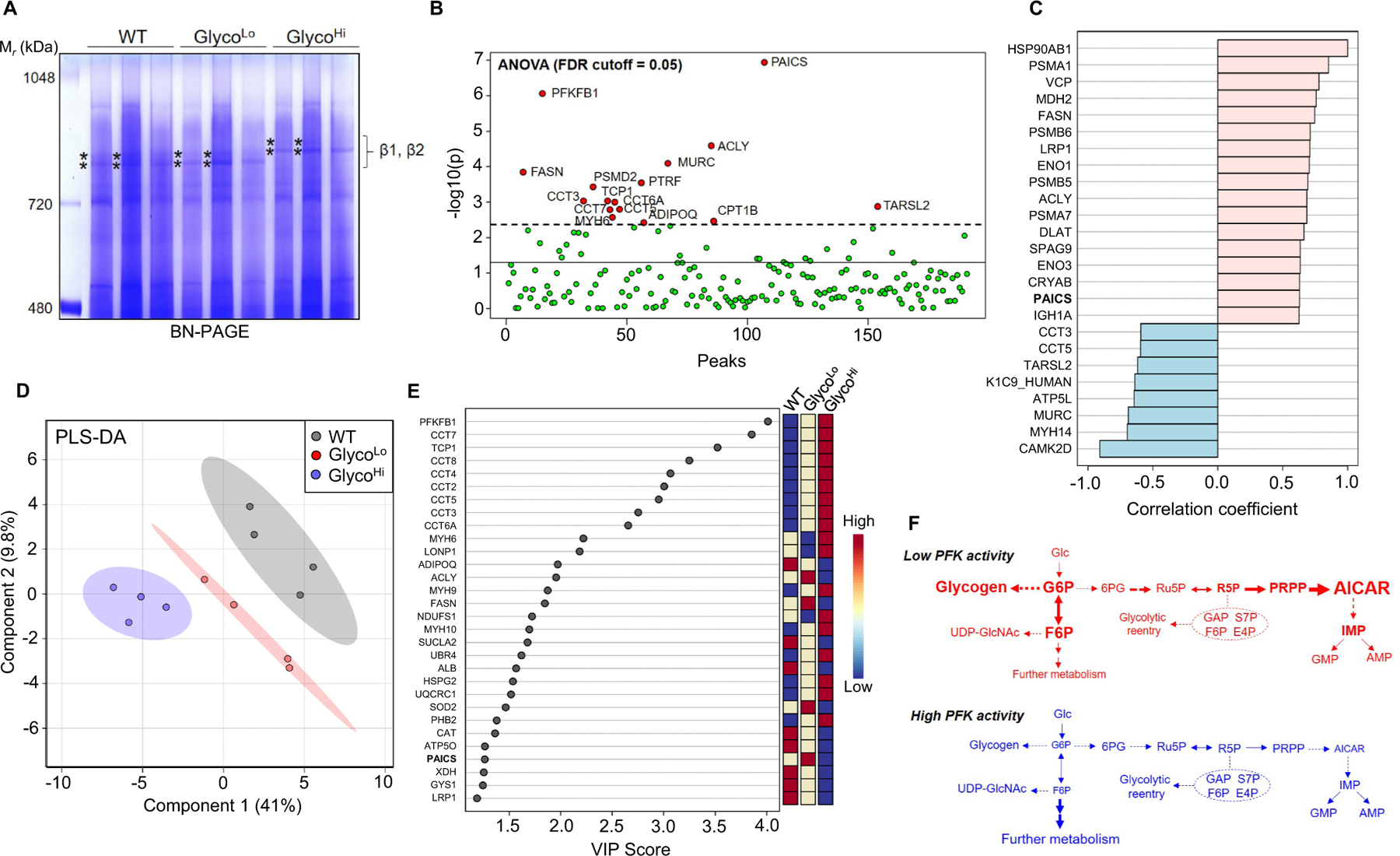
Glucose metabolism comprises numerous amphibolic metabolites that provide precursors for not only the synthesis of cellular building blocks but also for ATP production. In this study, we tested how phosphofructokinase-1 (PFK1) activity controls the fate of glucose-derived carbon in murine hearts in vivo. PFK1 activity was regulated by cardiac-specific overexpression of kinase- or phosphatase-deficient 6-phosphofructo-2-kinase/fructose-2,6-bisphosphatase transgenes in mice (termed GlycoLo or GlycoHi mice, respectively). Dietary delivery of 13C6-glucose to these mice, followed by deep network metabolic tracing, revealed that low rates of PFK1 activity promote selective routing of glucose-derived carbon to the purine synthesis pathway to form 5-aminoimidazole-4-carboxamide ribonucleotide (AICAR). Consistent with a mechanism of physical channeling, we found multimeric protein complexes that contained phosphoribosylaminoimidazole carboxylase (PAICS)-an enzyme important for AICAR biosynthesis, as well as chaperone proteins such as Hsp90 and other metabolic enzymes. We also observed that PFK1 influenced glucose-derived carbon deposition in glycogen, but did not affect hexosamine biosynthetic pathway activity. These studies demonstrate the utility of deep network tracing to identify metabolic channeling and changes in biosynthetic pathway activity in the heart in vivo and present new potential mechanisms by which metabolic branchpoint reactions modulate biosynthetic pathways.
Keywords: Anabolism; Channeling; Glycolysis; Metabolomics; Metabolons; Stable isotope.
Copyright © 2021 The Authors. Published by Elsevier Ltd.. All rights reserved.
Conflict of interest statement
Declaration of Competing Interest
The authors declare no competing interests.
Figures
References
-
- Cluntun AA, Badolia R, Lettlova S, Parnell KM, Shankar TS, Diakos NA, Olson KA, Taleb I, Tatum SM, Berg JA, Cunningham CN, Van Ry T, Bott AJ, Krokidi AT, Fogarty S, Skedros S, Swiatek WI, Yu X, Luo B, Merx S, Navankasattusas S, Cox JE, Ducker GS, Holland WL, McKellar SH, Rutter J, Drakos SG, The pyruvate-lactate axis modulates cardiac hypertrophy and heart failure, Cell Metab 33 (3) (2020) 629–648. - PMC - PubMed
-
- Atherton HJ, Dodd MS, Heather LC, Schroeder MA, Griffin JL, Radda GK, Clarke K, Tyler DJ, Role of pyruvate dehydrogenase inhibition in the development of hypertrophy in the hyperthyroid rat heart: a combined magnetic resonance imaging and hyperpolarized magnetic resonance spectroscopy study, Circulation 123 (22) (2011) 2552–2561. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
