

TAZ maintains telomere length in TNBC cells by mediating Rad51C expression
- PMID: 34488828
- PMCID: PMC8422726
- DOI: 10.1186/s13058-021-01466-z
TAZ maintains telomere length in TNBC cells by mediating Rad51C expression
Abstract
Background: Telomere maintenance is crucial for the unlimited proliferation of cancer cells and essential for the "stemness" of multiple cancer cells. TAZ is more extensively expressed in triple negative breast cancers (TNBC) than in other types of breast cancers, and promotes proliferation, transformation and EMT of cancer cells. It was reported that TAZ renders breast cancer cells with cancer stem cell features. However, whether TAZ regulates telomeres is still unclear. In this study, we explored the roles of TAZ in the regulation of telomere maintenance in TNBC cells.
Methods: siRNA and shRNA was used to generate TAZ-depleted TNBC cell lines. qPCR and Southern analysis of terminal restriction fragments techniques were used to test telomere length. Co-immunoprecipitation, Western blotting, immunofluorescence, Luciferase reporter assay and Chromatin-IP were conducted to investigate the underlying mechanism.
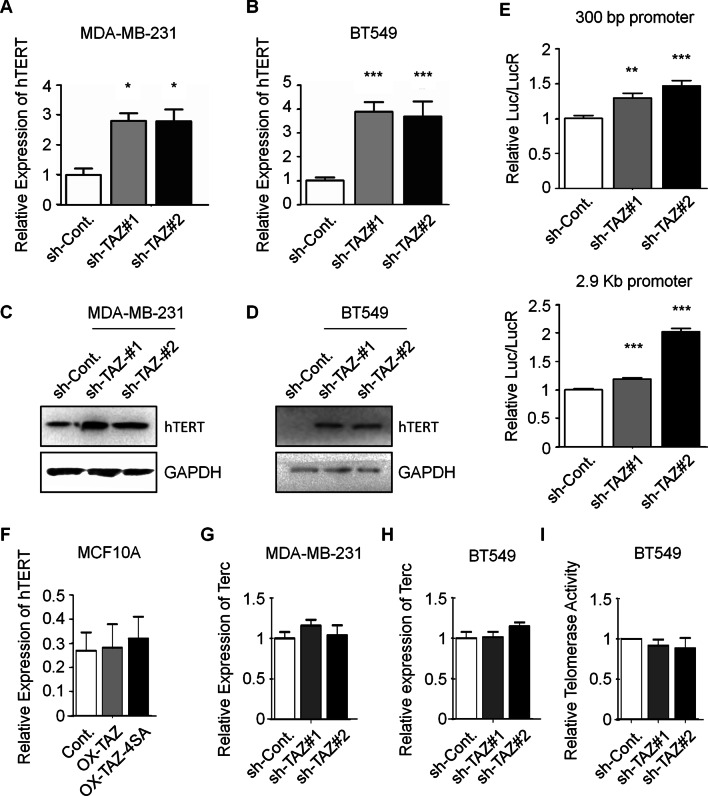
Results: By knocking down the expression of TAZ in TNBC cells, we found, for the first time, that TAZ is essential for the maintenance of telomeres in TNBC cells. Moreover, loss of TAZ causes senescence phenotype of TNBC cells. The observed extremely shortened telomeres in late passages of TAZ knocked down cells correlate with an elevated hTERT expression, reductions of shelterin proteins, and an activated DNA damage response pathway. Our data also showed that depletion of TAZ results in overexpression of TERRAs, which are a group of telomeric repeat-containing RNAs and regulate telomere length and integrity. Furthermore, we discovered that TAZ maintains telomere length of TNBC cells likely by facilitating the expression of Rad51C, a crucial element of homologous recombination pathway that promotes telomere replication.
Conclusions: This study supports the notion that TAZ is an oncogenic factor in TNBC, and further reveals a novel telomere-related pathway that is employed by TAZ to regulate TNBC.
Keywords: Rad51C; Shelterin proteins; TAZ; TERRAs; TNBC; Telomere.
© 2021. The Author(s).
Conflict of interest statement
The authors declare that they have no competing financial interests.
Figures






References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
