

Identification of a Novel Quinvirus in the Family Betaflexiviridae That Infects Winter Wheat
- PMID: 34489904
- PMCID: PMC8417474
- DOI: 10.3389/fmicb.2021.715545
Identification of a Novel Quinvirus in the Family Betaflexiviridae That Infects Winter Wheat
Abstract
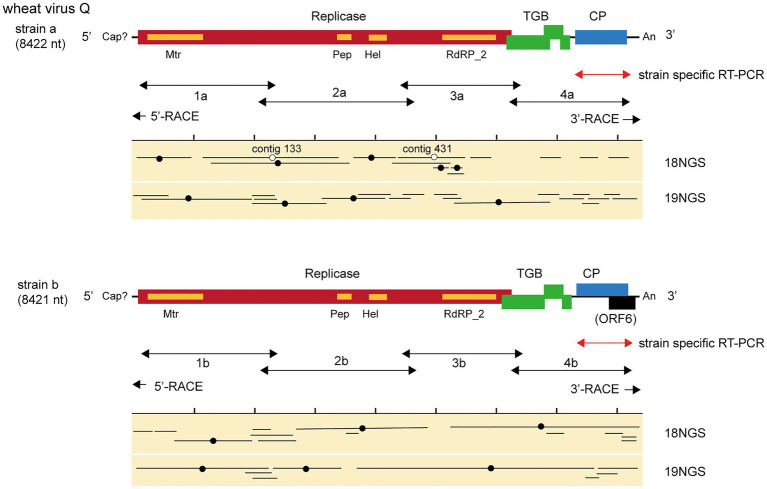
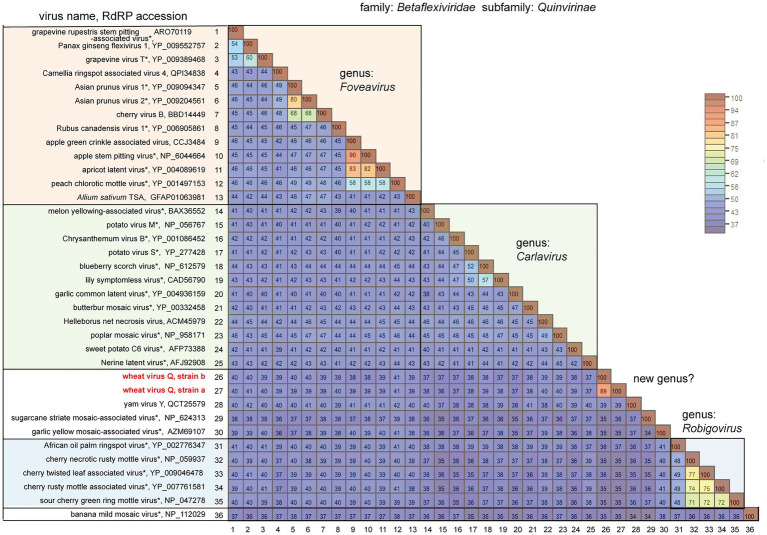
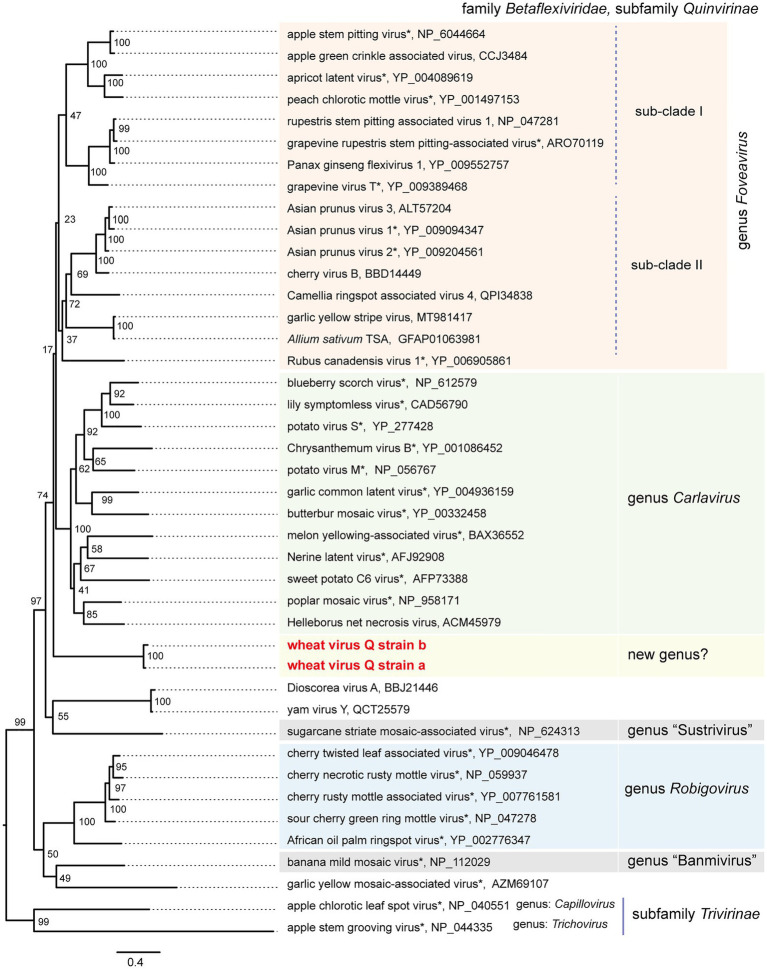
Yellow mosaic disease in winter wheat is usually attributed to the infection by bymoviruses or furoviruses; however, there is still limited information on whether other viral agents are also associated with this disease. To investigate the wheat viromes associated with yellow mosaic disease, we carried out de novo RNA sequencing (RNA-seq) analyses of symptomatic and asymptomatic wheat-leaf samples obtained from a field in Hokkaido, Japan, in 2018 and 2019. The analyses revealed the infection by a novel betaflexivirus, which tentatively named wheat virus Q (WVQ), together with wheat yellow mosaic virus (WYMV, a bymovirus) and northern cereal mosaic virus (a cytorhabdovirus). Basic local alignment search tool (BLAST) analyses showed that the WVQ strains (of which there are at least three) were related to the members of the genus Foveavirus in the subfamily Quinvirinae (family Betaflexiviridae). In the phylogenetic tree, they form a clade distant from that of the foveaviruses, suggesting that WVQ is a member of a novel genus in the Quinvirinae. Laboratory tests confirmed that WVQ, like WYMV, is potentially transmitted through the soil to wheat plants. WVQ was also found to infect rye plants grown in the same field. Moreover, WVQ-derived small interfering RNAs accumulated in the infected wheat plants, indicating that WVQ infection induces antiviral RNA silencing responses. Given its common coexistence with WYMV, the impact of WVQ infection on yellow mosaic disease in the field warrants detailed investigation.
Keywords: Betaflexiviridae; bymovirus; quinvirus; soil borne; variants; virome; wheat; yellow mosaic disease.
Copyright © 2021 Kondo, Yoshida, Fujita, Maruyama, Hyodo, Hisano, Tamada, Andika and Suzuki.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures



Similar articles
-
Differential Characteristics of Viral siRNAs between Leaves and Roots of Wheat Plants Naturally Infected with Wheat Yellow Mosaic Virus, a Soil-Borne Virus.Front Microbiol. 2017 Sep 20;8:1802. doi: 10.3389/fmicb.2017.01802. eCollection 2017. Front Microbiol. 2017. PMID: 28979249 Free PMC article.
-
Molecular characterization of a novel wheat-infecting virus of the family Betaflexiviridae.Arch Virol. 2021 Oct;166(10):2875-2879. doi: 10.1007/s00705-021-05175-y. Epub 2021 Jul 23. Arch Virol. 2021. PMID: 34297223
-
Association of VPg and eIF4E in the host tropism at the cellular level of Barley yellow mosaic virus and Wheat yellow mosaic virus in the genus Bymovirus.Virology. 2015 Feb;476:159-167. doi: 10.1016/j.virol.2014.12.010. Epub 2014 Dec 25. Virology. 2015. PMID: 25543966
-
Soil-borne viruses affecting cereals: known for long but still a threat.Virus Res. 2009 May;141(2):174-83. doi: 10.1016/j.virusres.2008.05.019. Epub 2009 Jan 19. Virus Res. 2009. PMID: 19159654 Review.
-
Bymovirus-induced yellow mosaic diseases in barley and wheat: viruses, genetic resistances and functional aspects.Theor Appl Genet. 2020 May;133(5):1623-1640. doi: 10.1007/s00122-020-03555-7. Epub 2020 Feb 1. Theor Appl Genet. 2020. PMID: 32008056 Review.
Cited by
-
Genomic and biological characteristics of a novel leafhopper-transmitted marafivirus infecting Triticum aestivum.Arch Virol. 2024 Mar 22;169(4):80. doi: 10.1007/s00705-024-06011-9. Arch Virol. 2024. PMID: 38519825
-
Analysis of Wheat Virome in Korea Using Illumina and Oxford Nanopore Sequencing Platforms.Plants (Basel). 2023 Jun 19;12(12):2374. doi: 10.3390/plants12122374. Plants (Basel). 2023. PMID: 37375999 Free PMC article.
-
Molecular characterization of a novel virga-like virus associated with wheat.Arch Virol. 2022 Sep;167(9):1909-1913. doi: 10.1007/s00705-022-05473-z. Epub 2022 Jun 25. Arch Virol. 2022. PMID: 35752685
-
Discovery and Genome Characterization of a Closterovirus from Wheat Plants with Yellowing Leaf Symptoms in Japan.Pathogens. 2023 Feb 21;12(3):358. doi: 10.3390/pathogens12030358. Pathogens. 2023. PMID: 36986280 Free PMC article.
-
Translation of Plant RNA Viruses.Viruses. 2021 Dec 13;13(12):2499. doi: 10.3390/v13122499. Viruses. 2021. PMID: 34960768 Free PMC article. Review.
References
LinkOut - more resources
Full Text Sources
Research Materials